



ادْعُ إِلَىٰ سَبِيلِ رَبِّكَ
شٌبهات وردود

المكتبة المرئية

خَــيْـرُ جَــلـيـسٌ

المتواجدون الآن
حرس الحدود
مجتمع المبدعين
البحث
عرض المادة
الملحمة المستحيلة: إشكاليات الاستدلال بسجل الحفريات على التطور
مقدمة
مرة أخرى، نعود لنشارك أنصار التطور رحلتهم الشاقة تنقيبا عن الحلقات المفقودة، ونتابع واحدة من أكثر قصص التطور شهرة وغرابة، والتي تعد من الأحداث الكبرى في تاريخ التطور، نعود لنلقى نظرة مختصرة على أحد أهم فصول هذه القصة.
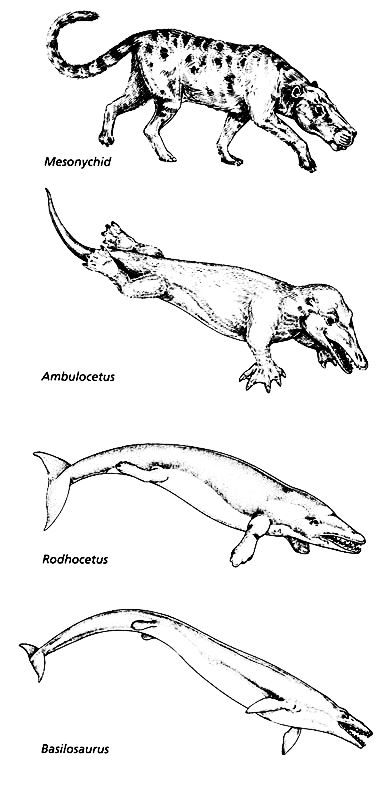
من خلال هذا الطرح نرصد تأريخ السجل الأحفوري لذلك الحدث الملحمي الذي يحكى لنا كيف تطورت وحوش اليابسة، التي كانت تسير على أربع، إلى وحوش البحار المهيمنة (الحيتان).

قصة تطور الحوت
الحيتانيات –الحيتان والدلافين– هي إحدى أضخم الثدييات، لكنها خلافا لمعظمها –التي تعيش على الأرض– تعيش حياتها كاملة في الماء، مع ذلك فهي ليست من الأسماك.
كان محتما على أنصار التطور حل تلك المعضلة، ووضع السيناريو الخاص بكيفية تطور هذه الثدييات وانتقالها إلى البحر مرة أخرى، بعدما غادرته قديما. لذلك افترض الطرح التطوري أن الحيتان ككل الثدييات تطورت من الزواحف، والتي تطورت بدورها عن البرمائيات، التي غادرت هي الأخرى المحيطات بعدما تطورت عن الأسماك، وذلك من حقب زمنية سابقة، حيث تركت الزواحف –أسلاف الثدييات– البحر منذ حقب زمنية سحيقة في تاريخ التطور، ونمت لها الأرجل، وكساها الفراء وتطورت الرئتان.
الأعجب هنا، هو إصرارها مرة أخرى على العودة إلى البحر، وذلك في حدث ملحمي استثنائي، لتفقد أرجلها وفرائها، وتبقي على رئتيها ونظام تكاثرها رغم هذه التحولات الجذرية التي طالت بنيتها.
ولدعم هذا الطرح، كان على أنصار التطور تقديم الأدلة والتنقيب في طبقات الأرض القديمة بحثا عن تسلسل انتقالي زمني بسجل أحفوري لأسلاف منقرضة كانت تسير على الأرجل، تظهر تدرجا وأشكالا وسيطة بين ثدييات الأرض وثدييات البحر، لكن ملامح ذلك السلف الأرضي القديم للحيتان ظلت شبحية برغم الافتراض السابق لداروين في مصنفاته –على استحياء– أنه كان دبا.

ولم تتكشف ملامح قصة تطور الحوت إلا بحلول عام 1966، حين قام عالم الأحافير فان فالين Van Valen أثناء فترة عمله بمتحف نيويورك للتاريخ الطبيعي برصد بعض التشابهات بين عظام مجموعة من آكلات اللحوم المنقرضة المسماة بوسطية الحوافر (الميزنيقيات Mesonychids) مع أحافير وعظام الحيتان Cetacea.
هذه الحيوانات المنقرضة الشبيهة بالذئاب والمسماة بـ(Mesonychids) امتلكت أسنان ثلاثية شبيهة بتلك التي في الحيتان المعاصرة، واستنتج فالين من تلك المشاهدة أن الحيتان انحدرت منها.(1)
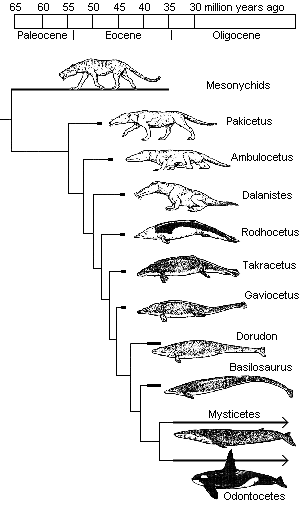
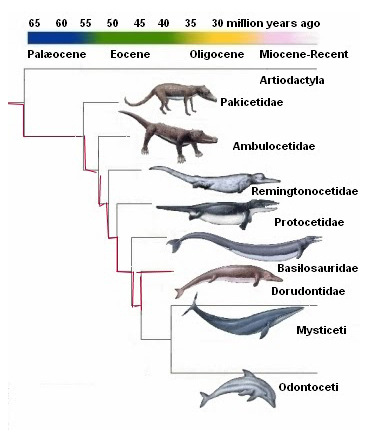
منذ ذلك الحين توجهت بوصلة أنصار التطور وعلماء الأحافير للبحث في ذلك الاتجاه الذي اقترحه فالين عن عظام أسلاف الحوت، التي تمتلك صفات مورفولوجية (هيكلية– تشريحية) مرتبطة بمجموعة وسطية الحوافر Mesonychids، وكان رائد البحث في هذا الاتجاه هو عالم الأحافير جنجريتش Gingerich، المتخصص ببحثه عن أسلاف الحوت، والذي بدأ رحلته في البحث خلال عقد السبعينيات من القرن المنصرم، واستمر لأكثر من عقدين في رسم الأطر العامة لسجل تطور الحيتان وتبعاته، وتزامنت معه كشوفات حفرية أخرى ودراسات متعددة لعلماء أخرين، والتي نتج عنها تأطير للخطوط العريضة لذلك المسار عبر سجل أحفوري مفترض، اصطفت خلاله سلسلة كاملة من الحيوانات المنقرضة واحدا بعد الآخر في تتابع زمني، وفقا للفترات الجيولوجية التي كانوا يعيشون فيها، ووصفت بأنها أشكال انتقالية متسلسلة بين الثدييات البرية والثدييات المائية بالكامل على نحو ما، كما يظهرها المخطط التالي:
|
قبل 50 مليون سنة |
باكيسيتوس Pakicetus الأرضية بالكامل |
| قبل 49 مليون سنة |
أمبيلوسيتوس Ambulocetus شبه المائية |
| قبل 46 مليون سنة |
رودهوكيتوس Rodhocetus شبه المائية |
|
قبل 37 مليون سنة |
باسيلوسورس Basilosaurus المائية بالكامل |
هكذا يحكى التطور قصة تحول الحوت بمخططات رائعة وقصة مثيرة، وهذا النوع من القصص يروقنا جميعا.
لكن دعونا نتعمق مع تلك القصة بجرعة أكبر من الإثارة، ونحاول إلقاء نظرة مدققة على هذا المقترح الذى يسجله السجل الأحفوري كعنصر وحيد للطرح، ولا ضير من أن نفتح أثناء عروجنا داخل أعماق تلك القصة أكثر من نافذة، نرى من خلالها مغالطات متأصلة وعامة في أصل المنهج التطوري، وقبل أن نبدأ بعرضنا هذا، أعدوا لأنفسكم فنجانا من القهوة وانعشوا ذاكرتكم ببعض التركيز لمتابعة تسلسل الأحداث.(2)
الإشكالية الأولى: هل سجل الحفريات دليل على التطور؟
عنوان الإشكالية السابق يطرح سؤالا بدهيًّا يتعلق باستعراض كفاءة الأحافير كدليل على السلف المشترك، بالتالي على صحة الاعتقاد في التطور الدارويني ككل، والإجابة عليه كفيلة وحدها بتحديد مصير الاستناد إلى تلك الدلالة.
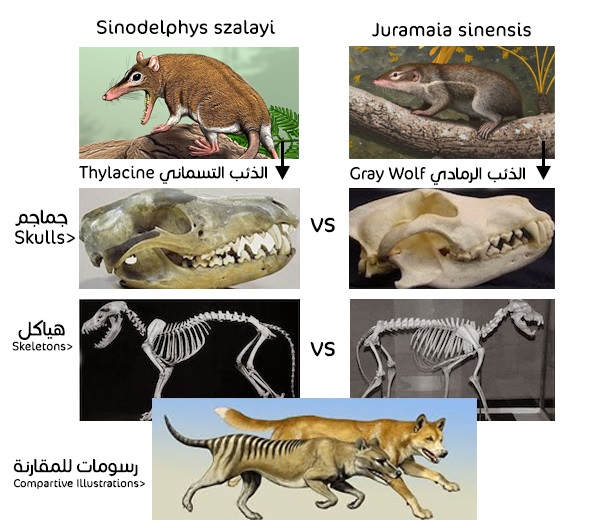
إنَّ الإجابة عن هذا السؤال يمكن توضيحها بمثال بسيط للغاية؛ لنفرض أننا وجدنا أحفورة لكائن ما، فإن الحقيقة الوحيدة التي نستطيع تأكيدها على وجه اليقين من خلال رصد تلك الأحفورة هي أنها كانت لكائن حي –مات ودفن– في هذا المكان، فيما عدا ذلك لا توجد أيّة وسيلة لدى أيٍّ من العلماء تؤكد يقينا أن هذه العظام التي وجدت مدفونة تمثل سلفا أو جدًّا لأى كائن حي آخر بمجرد رصد بعض التشابهات بينهما؛ لأنه فضلا عن استحالة تأكيد افتراضية السلف المزعوم تلك، فإن التشابهات المورفولوجية (التشريحية/الهيكلية) التي يُستند عليها في تأكيد مدى قرابة الأحافير لا تدل حتما على أي قرابة مزعومة، وهذه الإشكالية يدركها أنصار التطور جيدا، وإلا لأصبح هذا الذئب الأسترالي الجرابي قريبا من الدرجة الأولى لنظيره الذئب الرمادي المشيمي وفقا لهذا التطابق المذهل في المورفولوجي.
لكن الحقيقة أن الذئب الرمادي هو أقرب تطوريا للفيل والأرنب والإنسان، من هذا الذئب الجرابي، الذي يعتبر هو الآخر أكثر قرابة للكنغر والكوالا والسنجاب الأسترالي الطائر، عن مدى قرابته للذئب الرمادي التوأم التشريحي له. وذلك وفقا لما رسمته شجرة التطور؛ حيث افترضت انفصالا تطوريًّا بين أسلاف كل من الجرابيات والمشيميات في فجر نشوء الثدييات، منذ ما يقارب 160 مليون سنة مضت، وتكرر رصد مثل هذه التوائم المورفولوجية بين الجرابيات والمشيميات بنسبة عاتية، مقارنة بعدد الجرابيات المعدود على سطح كوكبنا.(3)
تلك التوائم المتماثلة التي لا يمكن ربطها بسلف مشترك –بمعايير التطور–لم تقتصر على فئتي الجرابيات والمشيميات فحسب، بل تم رصدها على كافة المستويات التصنيفية داخل الممالك الأحيائية، ورصدت أيضا تشابهات بالغة التعقيد على النطاق الجزيئي، كما تم رصدها على النطاق المورفولوجي.
وقد تناولت في طرح سابق جزءًا يسيرًا منها، متعلقًا بتوائم الجرابيات والمشيميات، والذي سنتبعه لاحقا –إن أذن الله تعالى– بمنشورات أخرى، نعرض فيها أمثلة كثيرة مدهشة ومتنوعة من التماثل المُعارض لشجرة التطور، تشمل كافة المستويات التصنيفية، مورفولوجيا وجزيئيا، وكيف كانت ردة فعل أنصار التطور في مقابل تلك الإشكالية، واعتراف البعض منهم ممَن واتتهم الجرأة على نقض الدوجما التطورية، والخروج خارج الإطار المعتاد، مما يمثل تهديدا لمصداقية شجرة التطور، وتداخل البيانات بين ما يعدونه تشابها ناتجا عن سلف مشترك، وآخر غير مرتبط بسلف.(4)

للخروج من المأزق
مثلت الإشكالية السابقة تحديا لأنصار التطور لا يمكنهم تجاوزه؛ ذلك لأنها تطعن مباشرة في أصل الاستدلال بالتشابه بين الأنواع على السلف المشترك، لأن ذلك التشابه (المورفولوجي والجزيئي) الذي يعد الدليل الأوحد على التطور يعطي دلالة معاكسة تماما، كما أظهرت الأمثلة الأخرى سالفة الذكر تماثلات خارج إطار السلف المشترك، لذا لجأ أنصار التطور إلى الالتفاف غير المباشر حول هذه الإشكالية بسلوك المنطق الدائري، والادعاء أن تلك التشابهات التي تتعارض مع قواعد شجرة الأنساب الافتراضية هي في الحقيقة غير مرتبطة بسلف تطوري، أي أن كلا النوعين اللذين أظهرا هذا التماثل قد سلكا طريقين منفصلين تماما في التاريخ التطوري لإنتاج نفس الهياكل المتماثلة أو نفس التشابه الجزيئي، وأطلقوا على هذه الفرضيةconvergent evolution ، كواحدة من فرضيات الخروج من المأزق. ما تم هنا في الحقيقة ما هو إلا إعادة توصيف لمشاهدة تطعن في أصل فرضية التطور، ولي عنق القاعدة –التي قعدوها لأنفسهم–، بل وكسره لتوافق البِرادايم Paradigm أو الإطار التطوري الذي لا يجوز الخروج عليه.
لكن الخطأ المنهجي في هذه الفرضية هو الالتجاء لمغالطة منطقية جلية؛ وهي المصادرة على المطلوب، بجعل المطلوب إثباته –أو النتيجة المرجو الوصول إليها (التطور)– ومقدماته أو إحداها –التي يجب الاستدلال عليها (التشابه)– شيئا واحدا. والمغالطة المنطقية تحصل هنا حينما يتم افتراض صحة النتيجة التي يراد البرهنة عليها في المقدمات سواء بشكل صريح أو ضمني، وحين يتم الاستدلال بالنتيجة المرجو الوصول إليها كحقيقة أولية لبناء هذا الافتراض.(5)
وبدلا من إعادة ذلك الاستقراء الناقص المبني على دلالة التشابه بين الكائنات كدليل حتمي على وجود سلف مشترك، بسبب وجود ما ينقضه من معطيات مختلفة لا تؤيد هذا الاستقراء، توجهوا مباشرة الى بناء فرضية convergent evolution اعتمادا على أن التطور حقيقة، وقد عرفنا تلك الحقيقة من التشابه بين الكائنات!
لاحظنا بوضوح تجلي المنطق الدائري Circular reasoning الذي لا ذيل له ولا رأس، ولا نستطيع التفريق فيه بين المقدمات والنتائج. فحين يكون من المفترض أن تتصدر التشابهات طرائق الاستدلال على صحة التطور والاشتراك في سلف مشترك، وتقدم على أنها من أكثر دلائل دعم التطور، فإنه يصبح كافيا جدا لإسقاط هذه الدلالة رصد تضاربا يناقضها، ويؤكد أن التشابه لا يدل على سلف مشترك. لكن بدلا من ذلك لجأ أنصار التطور إلى اختراع فرضية التطور المتقارب convergent evolution، التي بنوها على التسليم بصحة التطور.
لكن لا تلبس تلك الدائرة أن تنكسر، فيفاجئنا رونالد ويست Ronald R. West عالم الجيولوجيا التطوري الشهير، ويجرؤ على الإقرار المباشر بالأخطاء المنطقية الفجة تلك في الاستدلال بسجل الأحافير، قائلا:
"على عكس ما يكتبه معظم العلماء، فإن سجل الحفريات لا يدعم نظرية التطور الداروينية؛ لأنها تلك النظرية (بمختلف إصداراتها) هي التي نستخدمها لتفسير السجلات الحفرية. لذلك فنحن مدانون بالوقوع في استدلال دائري حينما نقول أن السجل الأحفوري يدعم هذه النظرية".(6)
ويمكنك عزيزي القارئ تلخيص المشهد السابق في كلمات بسيطة للغاية هي: التطور حقيقة، لأن التطور حقيقة!
هذا ما يقوله التطوريون بلسان حالهم، بل ربما يقره البعض بصيغة أكثر تحايلا حين يواجه مثل هذه الإشكاليات المنهجية فيقول: التطور حقيقة مثبتة مفروغ من صحتها، لكن البحث في آلياته وطرائقه مازال مستمرا ومفتوحا. لكن الإشكال هنا: كيف يثبت التطور أصلا بدون ثبوت فرضية السلف المشترك؟
بذلك المنطق الدائري الهزيل يتمكن أنصار التطور من تجاوز أي تضارب يعرض أمامهم، فتستخدم دلالات التطور حسب الحاجة، وتعطل أيضا حسب الحاجة، ليذكرونا بتلك الحكاية القديمة من عهود جاهلية العرب، حين يصنع الرجل أصناما ومجسمات من العجوة لألهته التي يعبدها، ويظل عليها عاكفا بالتضرع، حتى إذا جاع أكلها.
نخلص من التفصيل السابق بوضع الإطار العام للتعامل مع كافة الأمثلة التي يُستخدم فيها سجل الأحافير كدليل يثبت صحة التطور، وهو أن التشابه المورفولوجي البنيوي الذي تظهره بعض العظام الأحفورية مع عظام كائنات أخرى، سواء كانت حية أو منقرضة، لا يمكن تصديره كدليل على صلة قرابة حتمية أو أي تطور مزعوم من سلف قديم، فنفس التشابه نجده بين عظام كائنات حية لا يمكن إدراجها في إطار شجرة أنساب مشتركة كما أسلفنا الذكر في المثال السابق.
هذا إن تجاوزنا عدم قدرة سجل الحفريات على تقديم دعم حقيقي لفرضية التشابه من الأساس، فالفقر الحاد في تلك السجلات يجعل تقديم البيانات اللازمة للمقاربة المورفولوجية المطلوبة أمرًا في غاية الصعوبة، فلا تتوافر لدينا الأنسجة الرخوة، ولا غالبية أجزاء الجسد، ويمكننا ببساطه أن نوضح ذلك بمثال عملي حي:
لنفترض أن (الذئب التمساني الجرابي) و(الذئب الرمادي المشيمي) قد انقرضا منذ وقت طويل، قبل أن نتمكن من رؤيتهما أحياء، وتم اكتشاف أحافير عظمية خاصة بهما، فإنه مما لا شك فيه أن التعريف الخاص بالتطور سوف يؤكد وجود صلة قرابة وثيقة بينهما في شجرة التطور، بسبب هذا التطابق المذهل في تركيب الهياكل العظمية، لكن الهيكل العظمى رغم اكتماله لا يعطي أي تصور حقيقي لنظام التكاثر، سواء كان جرابي أو مشيمي، فكلاهما يمتلك نظام تكاثر خاص مختلف تمامًا عن الآخر، ولا توجد بينهما أيّة صلة!
لذلك فالأمر يزداد تراكبا وتعقيدا، فالإشكالية لا تنتهي عند إسقاط دلالة التشابه بين الحفريات كدعم للتطور بسبب وجود دلالات عكسية فحسب –كما بينا بأمثلة التماثل أو ما أطلق عليه التطور المتقارب كتعريف مضلل وموهم–، لكن تتعداه إلى الفقر الحاد في دعم الأحافير لتقديم بيانات يمكن الاستناد عليها لعقد هذه المقاربة المطلوبة من الأساس.
لنتخطى تلك الإشكالية، التي نعتبرها أيقونة في نقد الكثير من دلالات التطور، وندخل مباشرة إلى عرض أطروحة التطور الخاصة بموضوعنا حول قصة أسلاف الحوت.
ملخص ما سبق
1) الاستدلا الدائري وتناقض دلالات التشابه المورفولوجي يسقط حُجية الأحافير على التطور والسلف المشترك.
2) الفقر الحاد في البيانات التي يمكن أن تقدمها الحفريات في مقارنة الخلايا الرخوة التي لا يمكن تحجرها.
الإشكالية الثانية: هل تبدو النماذج الانتقالية المعروضة والرسوم تمثيلا حقيقيًّا للأحافير التي عثر عليها؟
الخطوة الأولى بعد العثور على عظام ما لأحفورة ما، هي توصيف هذه العظام بدقة، وذلك بواسطة فريق مختص، والعمل على ترميم وبناء شكل نهائي يمثل الحيوان الكامل، هذا إن توفرت العظام الكافية لذلك بشكل يضمن رسم صورة تخيلية يُظن أنها الأقرب إلى الواقع. فهل هذا ما تم حقا؟
للإجابة على تلك الإشكالية، لنرى كيف تعامل أنصار التطور مع الحفريات التي تم العثور عليها وكيفية ترميمها، وسننطلق من بداية تسلسل الحلقات الانتقالية كما بمخطط التطور من الجد الأول، حتى وصول أسلاف الحوت إلى الحياة المائية الكامل.
أولا: باكيسيتوس Pakicetus

في عام 1983، أثار فيليب جنجريتش ضجة إعلامية بزعمه اكتشاف حفرية لأحد الأسلاف الأولى للحيتان، والذي عرف باسم الحوت الباكستاني (باكيسيتوس Pakicetus). ادعى جنجريتش أنه كان حيوانًا وسيطًا بين حيوانات اليابسة والحيتان، وأنه الحلقة الانتقالية الأولى لهذا التحول.

احتفت مجلة العلوم Scince العلمية المرموقة بهذا الاكتشاف، وتصدرت أغلفتها رسومًا كاملة لذلك الحيوان، حيث يمتلك ساقين بهما أغشية كالزعانف، ويقوم بمطاردة الأسماك كحيوان بحري صياد.(7)
لكن إلى أي دليل استند جنجريتش في ترميم وصنع هذا المخطط الكامل؟ فكل ما يمكننا قوله هو أن الأدلة الأحفورية المتاحة لديه في هذا الوقت تألفت فقط من بعض شظايا الجمجمة؛ (جزء صغير في الجمجمة، وعدد قليل من الأسنان، وجزء صغير من الفك)، هذه هي المعطيات الحفرية التي ادعى من خلالها أنصار التطور أنها مثلت جد الحيتان في وقت مبكر في العصر الأيوسيني، وذلك بناءً على رصد بعض التشابهات في قمة الأسنان مماثلة لوسطية الحوافر المنقرضة Mesonychids –التي اعتقد فالين أن لها علاقة وثيقة بالحيتان الحديثة–، وبعض التشابهات في أجزاء من الجمجمة.
لكن بعد الفحص الدقيق للبيانات، لا زلنا نتساءل بدهشة لماذا تم اقتراح هذه العينة لأن تكون أي شيء. ظلت هذه الفكرة السائدة عن شكل باكيسيتوس لفترة؛ يحتفى بمخططه الكامل كما رسمه جينجريتش دون أن يُقدم أحد من أنصار التطور على الاعتراض البديهي السابق. لكن بحلول عام 2001، قام الخبير البارز بأحافير الحيتان هانز ثيوسين J. G. M. Hans Thewissen صديق جينجريتش وزميله حسين وفريقهما بإعادة إعمار أكثر حداثة، اعتمدوا فيها على اكتشافات لأكثر عظام الحيوان باكيسيتوس، ونشرت مجلة نيتشر ذلك الكشف.
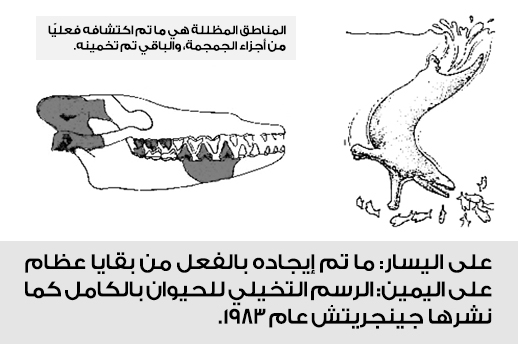
مع إعادة ترميم أكثر معقولية تمت على تلك العظام وبعد إيجاد حفريات جديدة له، كانت المفاجأة أن المخطط الجديد كان لحيوان بري كامل لا يشبه الأول في شيء.
وأكد ثيوسين في نفس الورقة إلى أن تلك العظام تشير بوضوح إلى ثدييات برية بالكامل، بل وتظهر السمات التشريحية لعظام تحت القحف أن الحيوان كان من العدائين، ولم تلامس أقدامه إلا اليابسة.(8)
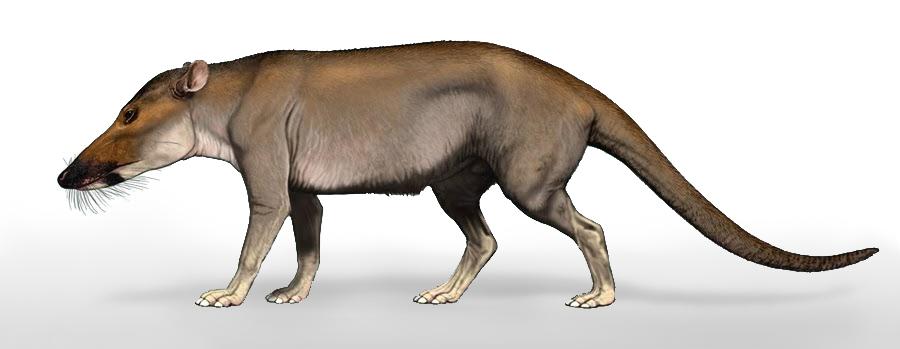
ربما نلتمس عذرا لجينجرتش في التجائه لهذه الرسوم المضللة في عام 1983، بسبب إحباطه وملله من البحث عن شيء ذي قيمة لسنوات عديدة، فكانت بعض شظايا الجمجمة التي تبدي بعض التشابه مع عظام الحوت كفيلة بصنع أسطورة الحوت الأول الصياد، لذلك لجأ إلى الخيال، ولم يتورع عن فعل ذلك؛ لأنها كانت سمة ظاهرة في تحليل أنصار التطور لعظام الأحافير، ومثل هذه الحوادث متكررة في كثير من المخططات الموهمة المتداولة، التي تم بناؤها على حفريات غير مكتملة، لكن هناك نوعًا آخرَ من التزييف الخفي المتعمد، التجأت له دوريّات حديثة، حتى بعد إعادة إعمار أكثر حيادية 
للأحفورة، مثلما فعلته المجلة العريقة والأكثر شعبية (ناشيونال جيوجرافيك National Geographic) في تصويرها لمخططات الحفريات بعد إعادة إعمارها.(9)
حيث نجد في المخطط نوعا من الإيحاء والإيهام المتعمد لتمرير فكرة سلف الحوت، فالتجأ الناشر إلى تدليس الفنان باستخدام صورة لـباكيسيتوس في وضع السباحة، مع ذيل مكتنز انسيابي كذيل السمكة، وأرجل قصيرة نسبيّا، رغم أنه حيوان بري كامل، ويلاحظ أيضا أن الرسوم تظهر الساقين الخلفيتين ممددتين إلى الوراء؛ لتعطي انطباعا أنها تعمل مثل الزعانف.
ثانيًا: أمبيلوسيتوس Ambulocetus

الحفرية الثانية تدعى أمبيلوسيتوس Ambulocetus، التي وجدت عام 1993 في باكستان، وادعى مكتشفه ثيوسين وآخرون أن هذه العظام كانت لأحد أسلاف الحوت، في حجم ذكر أسد البحر، الذي كان يمشي على أرجل، وكانت أرجله الخلفية بمثابة الأرجل على الأرض، والمجاديف –الزعانف– في المياه، مما يشير للاعتقاد بأن هذا المخلوق قد تمكن من المشي على الأرض، وكذلك السباحة. مرة أخرى تدعى حاجة أنصار التطور لإثبات نظرية تطور الحوت إلى نشر ورقة أقل ما توصف به أنها تخمينية بامتياز، لكنها مرّت على هيئة التحكيم دون الإشارة إلى عوز تلك الورقة إلى أي استنتاج خارج إطار التخمين غير المعتمد على أية معطيات فعلية.(10)
لنختبر حقيقة القصة!
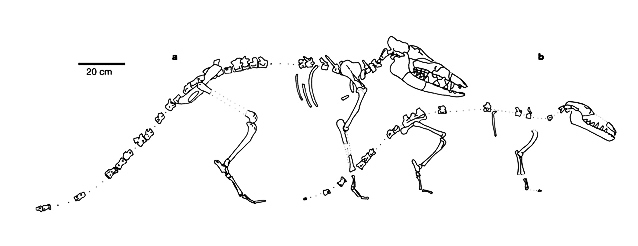
بالنظر إلى العظام التي وجدت لهذا الكائن المنقرض، فإن أول ملاحظة يمكن رصدها مباشرة في الكشوف الأولية هي أن الهيكل العظمي أيضا غير مكتمل، ويفتقد الأجزاء الهامة والجوهرية التي بُني عليها هذا الرسم التخطيطي للكائن البرمائي، والتي لا يمكن تصميم أي مخطط بدونها؛ لأنه يلزم لإنشاء وظيفة الساق الخلفية وترميمها تواجد عظام منطقة الحزام الحوضي pelvic girdle، لكنها مفقودة تماما في العظام التي عثر عليها.
وتم الترويج لهذه الرسوم التخيلية أيضا في كتب التدريس رفيعة المستوى، من قبل الأكاديمية الوطنية للعلوم. لاحظ الخيال المستخدم في الرسم، بما في ذلك أغشية وشبكات بين الأقدام.(11)
حتى (كينيث ميلر) كان له أيضا نصيب في نشر هذا التزييف بصورة مباشرة، حين استخدم هذه الحفرية كأيقونة مهمة للتطور في كتابه Finding Darwin's God، حيث زعم أن هذا الحيوان يمكن أن يتحرك بسهولة سواء في البر أو في الماء.
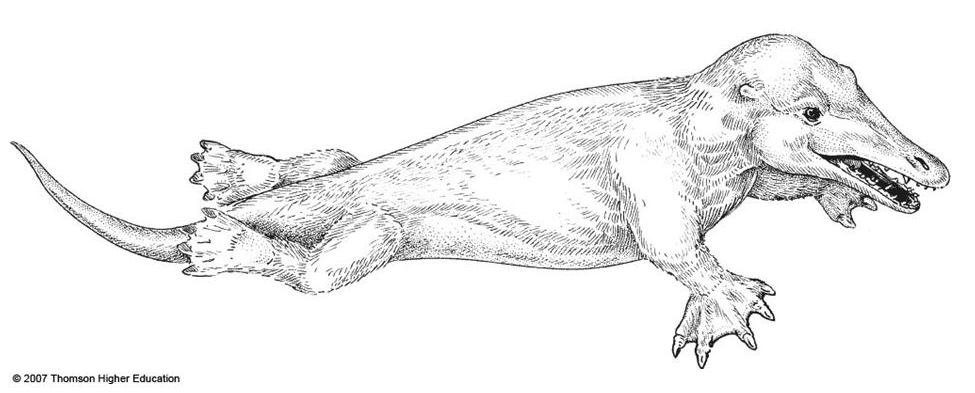
وكالعادة، (ناشونال جيوجرافيك) كان لها نصيبها المماثل من التلاعب والخداع البصري في تصميم مخطط الأحفورة، بإيحاءات بصرية تم توظيفها بعناية؛ لتحويل أمبيلوسيتوس البري إلي حيوان سباح، حيث أظهرت شبكات وهمية بين مخالب الحيوان، تمامًا كما فعلت دورية الأكاديمية الوطنية للعلوم و(كينيث ميلر) في كتابه، وجعلت الأرجل الخلفية تبدو وكأنها أرجل كسيحة على الأرض لا يمكن أن تساعده على المشي، بل تعمل كزعانف.(12)

وبالرغم من تأكيد (كارول) في إعادة التعمير –الأكثر معقولية وقبولًا– أنه كان حيوانا يمتلك أرجلًا قوية يستخدمها في المشي، لكن يبدو أن أنصار التطور يبحثون عن أي سبيل لتأكيد فكرة الحلقات الوسيطة، ولو بالتحايل والخداع البصري.(13)
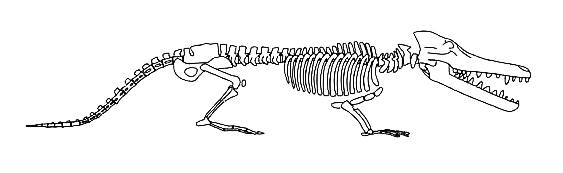
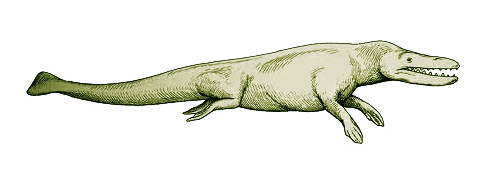
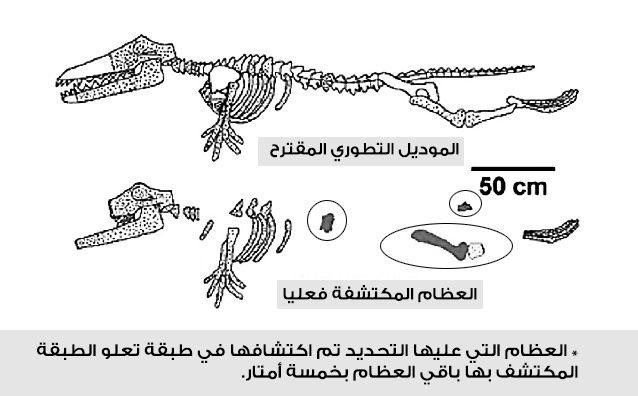
ثالثا: رودهوكيتوس Rodhocetus
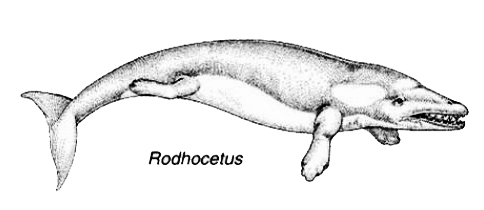
المرشح الثالث حسب الترتيب الزمنى للحلقات الوسيطة بين الثدييات البرية والحوت هو رودهوكيتوس Rodhocetus، ويُصوّر في المتاحف والكتب المدرسية كمخلوق لديه ميزات جوهرية للتحول من حيوان بري إلى حيوان بحري؛ حيث لوحظ تشكل الساقين إلى ما يشبه الزعانف، ونمو الذيل الشبيه بذيل الحوت.
|
مخطط: الرسوم التوضيحية للأكاديمية الوطنية للعلوم NAS. |
أثناء تصوير الفيلم الوثائقي عن التطور (تجربة عظيمة)، لاحظ الدكتور كارل فيرنر، القائم على التوثيق، تباين العرض الأحفوري للحيوان بجامعة ميشيغان والحفريات الفعلية، خاصة مع عدم وجود أيّة أحافير تظهر الذيل أو الزعانف، وهي الأشياء ذاتها التي يتم استخدامها كدليل على أن هذا المخلوق هو الحلقة المفقودة في تطور الحيتان.
ولمناقشة تلك الإشكالية، قام (فيرنر) بإجراء مقابلة مع جنجريتش العالم المسئول عن اكتشاف وإعادة بناء رودهوكيتوس، وكان الدكتور جنجريتش قد روج لفكرة أن رودهوكيتوس كان يمتلك ذيلًا بدائيًّا للحوت. كانت المفاجأة في رده على سؤال فيرنر حول كيفية تخمين ذلك النوع من الذيل بدون وجود عظام تدعم الفكرة، بتأكيده أنها مجرد تكهنات.(14) بل اعترف جنجريتش أيضًا أن الزعانف قد تم تخمينها، دون وجود أدلة داعمة من العظام.(15)
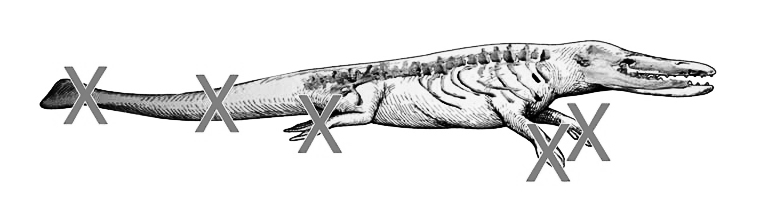
ونشاهد في الدقيقة (7:40) من الفيديو جنجريتش وهو يعترف أن الصور المتعلقة برودهوكيتوس –مثل الذيل– هي صورة متخيلة.(16)
الحقيقة أن هذه هي العظام التي تم العثور عليها كما يظهرها المرجع الشهير (كامبل).(17)
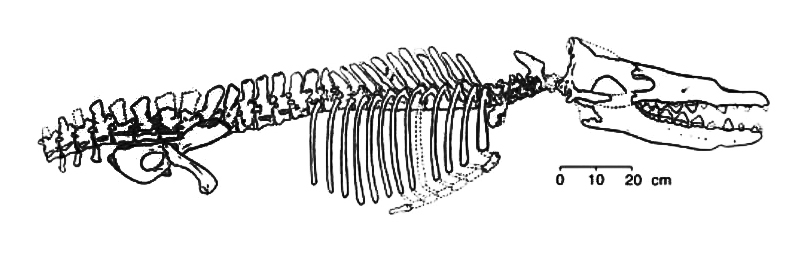
المزيد من العروض المضللة للأسلاف المزعومة للحوت أيضًا، ما نشرته الأكاديمية الوطنية للعلوم NAS من مخطط يوضح تسلسل التطور للحيتان، والذي يتضمن:Mesonychid التي تعيش في الأرض، يليها أمبيلوسيتوس و رودهوكيتوس وباسيلوسورس Basilosaurus، على الرغم من أن تقديرات المؤلفات العلمية تقول إن أطوال كل من أمبيلوسيتوس ورودهوكيتوس حوالي سبعة إلى تسعة أقدام، مقابل طول يقدر بحوالي 70 قدما في باسيلوسورس، إلا أن كتيب NAS أظهر جميع العينات الأربع على أنها بنفس الحجم، دون أن ينوه حتى في الحاشية –للتوضيح– أنه قد تم اختيارهم من الحجم الكبير! ومثل هذه الأخطاء لا يقع فيها مبتدئ في دراسة وعرض الأحافير، مما يثير الشك في كونها تدليسًا متعمدًا.(18)
تابعوا معنا فيديو الدكتور (تيري مورتنسن)، يبين فيه جانبا من مشاكل ترميم الأحافير السابقة، ويعرض مفاجآت وسط استغراب الحاضرين.(19)
ملخص ما سبق:
لجأ أنصار التطور إلى نوع من التضليل في بناء المخططات الكاملة لأنواع منقرضة، بناءً على ما تمّ العثور عليه من بعض عظامها، وذلك لتبدو أشبه بحلقة وسيطة بين أنواع مختلفة؛ لدعم أفكارهم.
وكما هو معهود، فإن خيال الفنان الموجه هو اللاعب الأساس في ترميم ورسم المخططات الكاملة لأغلب الحفريات، لتبدو للمشاهد كأنها تمثل حلقة وسيطة فعلًا، ودون اعتماد أية معطيات علمية حقيقية داعمة لهذه الرسوم المنشورة، وبمجرد أن ننظر نظرة متأنية في سجل أحافير الحوت سندرك هذه الحقيقة للأسف، إذ لا بأس عندهم من وضع عشرات المؤثرات البصرية لترسيخ الفكرة؛ فهنا جانب من مغامرات الصيد:

وهنا قليل من حنان الأمومة:

وهكذا، حتى نقبل الوهم ونصدقه!
لكن دعونا نتخطى ذلك، ونفترض أن تلك الأحافير كانت سليمة وكاملة حقا، فهل يمكن أن تدل على علاقة تطورية فعلية؟
الإشكالية الثالثة: السجل الأحفوري ومتاهة (مكعبات الليجو)
في الرسم البياني المقابل لسجل تطور الحيتان: أمسكْ قلمًا، وتتبع معي من نقطة البداية عند أي نوع من الأنواع الموجودة، وتحرك منه على خط السلف نزولًا بالزمن، حتى تصل إلى أصل هذا النوع ونهاية التفرع بالعودة إلى الماضي.
هل لاحظت شيئا؟
نعم، أنت لم تقابل أثناء مد الخط وفي رحلتك الزمنية نحو الماضي أي سلف أو جَدّ مباشر لهذا النوع.
جرب مع نوع آخر؛ مازالت نفس الإشكالية!
جرب مع كل الأنواع؛ الإشكالية نفسها!
إذن، أين هي الحلقات الوسيطة؟ وأين أسلاف الحوت؟
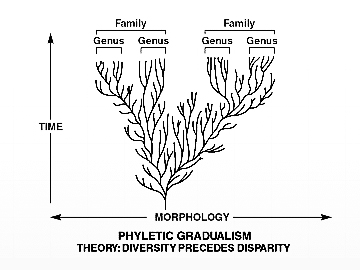
قبل الخوض في أي شيء يخص هذه الإشكالية، يجب علينا أولًا أن نعي جيدًا أن التطور ببساطة هو التغيير التدريجي من نوع إلى آخر خلال مدى زمني معين كما تقدمه لنا الداروينية.
نموذج الداروينية الحديثة يرسم توقعًا بوجود الكثير من الكائنات الانتقالية التي تمثل التغيرات التدريجية الصغيرة الناجمة عن طفرات جينية عشوائية التصرف يعمل عليها الانتقاء الطبيعي.
من خلال هذا التعريف، فإن السجل الأحفوري يُعنى بتوثيق العلاقة التطورية بين نوع معاصر وسلفه أو جده المفترض بحلقات وسيطة لأشكال انتقالية، يمكنها الربط المباشر بينهما كتدليل نظري على صحة هذا الفرض. أي أن هذا الخط الزمني بين النوعين يجب أن تتخلله وتتراص بداخله آلاف من الأشكال الانتقالية المتدرجة مورفولوجيا، ومن خلال هذا السريان التدريجي وعدم الانقطاع يمكن التأسيس لما يسمى بالسجل الأحفوري لتحول مفترض –نظريّا–، حتى يمكننا البدء في قبول تلك الفكرة مبدئيًّا كمشاهدة يمكن استقراؤها على نحو ما.
لكننا نلاحظ في السجل الأحفوري للحوت –كغيره من سجلات الأحافير– أن الأشكال الانتقالية التي يمكنها رسم أي علاقة تطورية بشكل تدريجي ليست متوافرة لتدلي بأية معلومة يمكن الاعتماد عليها في رسم تنسيق معين وخط انتقالي تصاعدي منضبط وثابت، ينتقل عبر الطبقات الجيولوجية والزمن، ويشير إلى هذه العلاقة التطورية المزعومة بين تلك الأحافير.
بذلك فإن نموذج "الداروينية الحديثة" يواجه العديد من الصعوبات العلمية، وأهم المشاكل التي تواجهه هي أن السجل الأحفوري بالكامل متقطع للغاية، مع غياب ساحق للأشكال الانتقالية بين جميع الأنواع الرئيسة. والواضح تمامًا أن السجل التاريخي للحياة –فعليّا– لا يوفر الأدلة اللازمة لتغيير تطوري تدريجي.
الحقيقة السابقة حول الفقر المدقع لسجل الأحافير لا يجادل فيها أي باحث مختص، وقد أشار أكثر علماء التطور إلى تلك الإشكالية، بداية من (داروين) نفسه وحتى يومنا هذا، رغم أن لدينا الآن أكثر من مائة مليون أحفورة في المتاحف المختلفة، بالإضافة لتلك الموجودة بالجامعات وجهات الأبحاث، وكلها تمثل أكثر من 250 ألف نوع مختلف.(20)(21)(22)
بنظرة أولية لسجل أحافير الحوت، يمكننا رصد غياب تام لأيّة أحفورة تمثل الحلقة الوسيطة أو الجَدّ المشترك بين أي من الأسلاف المزعومة، بل يظهر بوضوح مدى زمني ظهرت به مجموعة من الأحافير لأنواع معينة لم تتغير أو يطرأ عليها أي تطور خلاله.
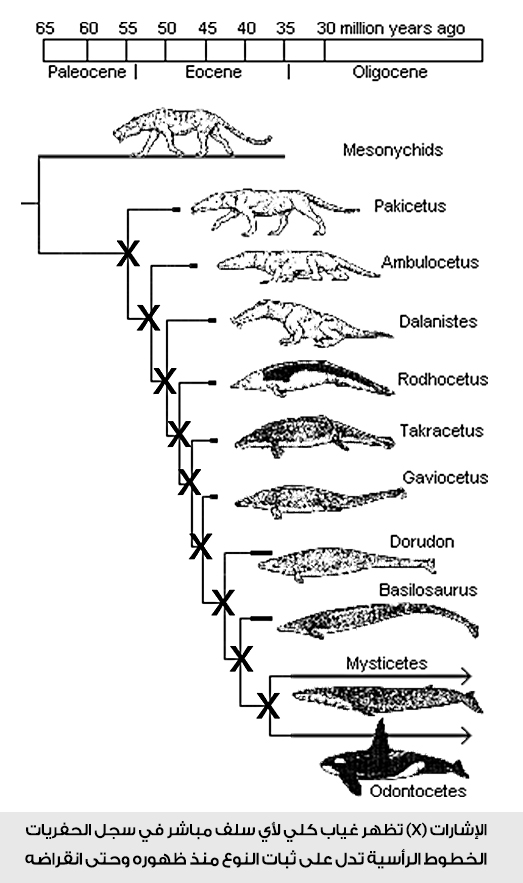
أي أن كل الحفريات كانت لأنواع ظهرت فجأة في السجل الجيولوجي، وعاشت في حقب زمنية لم يطرأ عليها خلالها أي تغيير، إلى أن انقرضت أو استمرت كما هي حتى الآن، كما في حالة الحوتيات الحديثة المعاصرة.
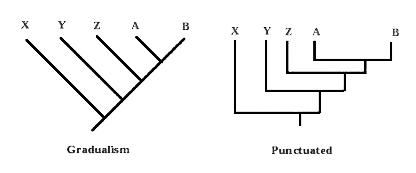
المخطط البياني التالي لسجل حفري، كما يجب أن يتم رصده بحسب (الداروينية الحديثة)، وتدعمه الآلاف أو الملايين من الحلقات التي يمكن عن طريق تسلسلها رسم منحنى تدريجي بتغير انسيابي سلس، للانتقال بين الأنواع بتفرع شجري معروف.(23)

شاهد المخطط البياني الفعلي للسجل الأحفوري المرصود، فهو يُظهِر خطوطًا عمودية للانتقال الزمني للنوع دون أي انحناء، مما يشير إلى امتناع التغير المورفولوجي والنوعي بالمطلق، ويشير إلى الظهور المفاجئ للنوع داخل السجل الجيولوجي، وبقائه ثابتا حتى انقراضه. وشاهد الربط الأفقي المباشر بين الأنواع، للإيحاء بالقرابة بين النوعين داخل السجل الأحفوري المرصود، لكن الخط الأفقي تمامًا الرابط بين نوعين يؤكد على حقيقة الظهور المستقل لكلا النوعين دون العثور على أي سلف مشترك بينها، ودون أدنى تدرج تدعمه حلقات وسيطة يمكنها أن تلعب دورَ قطع (الليجو) أو الفسيفساء بينهما.
هذه المشاهدات لا تقتصر على سجل أحافير الحوت، لكنها إشكالية عامة على كافة المستويات التصنيفية بشجرة التطور، وكل فروعها المرسومة، كما يقر بذلك المختصون من أنصار التطور في العديد من المواضع.(24) (25) (26) (27) (28)
هذه الحقيقة لم يتسنّ لأنصار التطور إخفاؤها، لكن أمكنهم التحايل عليها بالفرضيات المضللة والدائرية، وذلك باعتبار هذه الأحافير أبناء عمومة وليست أسلافا مباشرة لبعضها، وتم تسكينها داخل أفرع تطورية جانبية، دون توظيفها كجد حقيقي للأنواع الحالية، أي أنهم اعتبروا أن هذه الحفريات تطورت هي الأخرى من السلف المشترك الذي لازال مفقودا، وليست هي السلف الحقيقي، بذلك تكون قد سلكت فرعًا تطوريًّا مستقلا.
هكذا يمكن لأنصار التطور الهروب بذكاء من أيّة مسائلات أو إلزامات علمية حقيقية حول فقر السجل الأحفوري وعوزه إلى آلاف الحلقات الانتقالية، لنستطيع أن نصنع فروعَ شجرة الأنساب، التي لا يمكن اعتمادها نظريًّا إلا بوجود هذه الآلاف أو الملايين من الحلقات، تمثلها الأحافير المتدرجة مورفولوجيا، وتسكينها وفق منهجية صارمة؛ لتسير في خط تطوري تدريجي نحو التطور للنوع الجديد، وتتسق تتابعاتها صانعة انحناءً بيانيًّا زمنيًّا واضحا في رسوم شجرة الأنساب، يمثل فروع تلك الشجرة المزعومة، وهو ما لم يتوفر أبدًا في السجل الأحفوري للحوت، فضلًا عن أي من سجلات الأحافير المتاحة لأي نوع آخر، حيث تعاني كل السجلات الأحفورية الفقر الشديد في عدد الأحافير التي يمكن توظيفها كدعم لفكرة (التطور التدريجي) المزعوم، ورسمة شجرة الأنساب المتعلقة به.
لكن كما يؤكد أكثر العلماء المختصين، فإن فرص نجاح البحث عن تلك الأشكال الانتقالية المزعومة التي تمثل الأسلاف هي مثل فرص البحث عن سراب؛ ذلك لأن فقدها ليس متعلقًا بإمكانات البحث أو قلة عدد الأحافير التي عثر عليها حتى الآن، بل يرجع لكون السجل الأحفوري يمثل بالفعل الأحداث الحقيقية للحياة.(29)(30)
الظهور المفاجئ للأحافير –كما يظهرها السجل الأحفوري– يعني عدم التدرج السلس في التحول، لكن السمة الاكثر تحديًا للنموذج التطوري هي الثبات أو (الركود Stasis)؛ حيث تظل الأحافير للأنواع المختلفة ثابتة مورفولوجيًّا، دون أدنى تطور تظهره طبقات جيولوجية متعاقبة، وهذا ما يعبر عنه في الرسم البياني للسجل الأحفوري بالانتقال العمودي عبر الزمن –كما سلف ذكره–، وما يمكن معاينته من خلال السجل الأحفوري كما عرفه العلماء.(31)(32)
للخروج من المأزق
عدم وجود دعم من السجل الأحفوري، وندرة الأشكال الانتقالية التي يمكنها أن تؤلف الحد الأدنى المعقول لرسم سلسلة متصلة من التغيرات المتدرجة لدعم التطور، سبّبَا حرجًا بالغًا للفرضية التطورية، مما أدى بهم إلى اللجوء لحيلة دائرية جديدة للهروب من المأزق، وهي التي يعتمدون عليها في ترقيع الفرضية التطورية، شبيهة تمامًا بحيلة Convergent Evolution من حيث اعتماد المنطق الدائري بالالتفاف والمصادرة على المطلوب، حيث بدلًا من الاستدلال بالتغير المتدرج على صحة التطور –واعتبار العكس دليلًا ناقضًا له–، تم تبرير الظهور المفاجئ والركود في سجل الأحافير داخل البرادايم التطوري من منطلق الدوجما التطورية كحقيقة مطلقة، فتم التأسيس لما يعرف بفرضية Punctuated equilibrium أو التوازن المتقطع، وهي التي تفترض أن التطور لا يحدث في خطوات تدريجية صغيرة، بل يحدث في قفزات قصيرة من الزمن، قبل التوصل إلى الاستقرار والركود.
هذا يعني أن الأشكال الانتقالية ستكون قليلة العدد، لأنها لا توجد إلا في المجموعات الصغيرة لفترات صغيرة من الوقت، لذلك كانت الفرصة ضئيلة لترك الحفريات، بالتالي فإننا لا نتوقع العثور على الأشكال الانتقالية أو الحلقات المفقودة.(33)
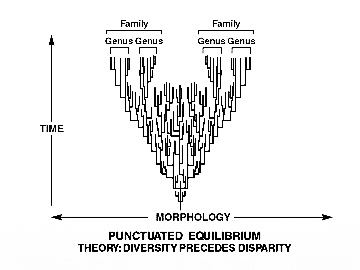
هذه الفرضية تواجه إشكاليات منهجية حقيقية كثيرة لازالت مثارًا للجدل، منها تعارضها هي الأخرى مع بيانات السجل الأحفوري، وإن كانت اعترفت بجزء من إشكاليات السجل الأحفوري من الانقطاع المتمثل في الظهور المفاجئ للأنواع، والثبات دون تغيير، والتفّتْ حولها بمنطق دائري، لكن يبقى ظهور تحت الممالك الرئيسة للأحياء Phyla كلها بانفجار نوعى هائل، حائلًا دون قبولها.(34)(35)
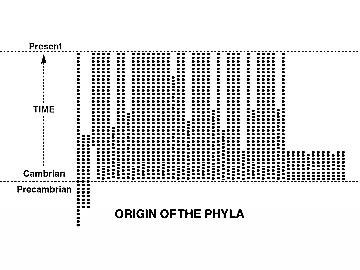
لكننا هنا نكتفي برفضها منطقيًّا؛ لأنها استخدمت التطور كمقدمة ومعطاة أولية للبناء، في حين يجب أن يكون التطور نتيجة واستقراءً، يستدل على صحته بالسجل الأحفوري، لذلك ينطبق بجدارة عليها اعتراض (رونالد ويست Ronald R. West)ـ.(36)
أما من حيث المنهجية التطورية، فإنها فرضية عادلة بما فيه الكفاية؛ ليحصل أنصار التطور على السلام النفسي بعد عناء من البحث المضني عن تلك الأشكال الوسيطة، التي سبّبَ غيابها لهم صداعًا لا ينقطع، لكن في المقابل، فإن الخطأ كل الخطأ هو أن يجرأ الآخرون على ادعاء أن السجل الأحفوري –بظهور الأنواع من خلاله فجأة وثباتها حتى الانقراض– هو دليل مباشر على الخلق المستقل للأنواع، وإلا تم اتهامه ومحاكمته بالهرطقة.
لكن من الإنصاف عدم التعميم، فهناك مَن يجرؤ دائمًا من أنصار التطور على تحدي هذه الدوجما صراحة أو اضطرارًا، فيعترف (مارك بريدلى) الذي يشغل حاليا منصب أستاذ علم الحيوان في جامعة أكسفورد بمثل هذا الخطأ المنطقي، وعدم صلاحية الاستدلال بالسجل الأحفوري، سواء كان ممثلًا بنموذج النيوداروينية المتدرجة gradualist، أو بنموذج التوازن المفاجئ punctuationist على التطور في مقابل الخلق الخاص، لأنه يدرك رجوح كفة الخلق الخاص المباشر في هذه المقاربة.(37)
متاهة الليجو

عودة إلى ألعاب الطفولة، وإلى (مكعبات الليجو) الشهيرة، التي طالما أثارت شغفنا عندما كنا صغارًا. حيث مكمن الإثارة في هذه اللعبة هو وجود مساحات شاسعة من الاختيار في تركيب وصنع عشرات المجسمات المختلفة من نفس المجموعة من قطع الليجو.
تخيل أن لديك قطعتين من اللعبة على مسافة متباعدة، نريد سد الفراغ بينهما بقطع أخرى، فإنه من البدهيّ لدينا أن مساحة الاختيار تضيق بضيق هذه المساحة الفارغة بين هاتين القطعتين؛ فكلما ضاقت المسافة كان عدد قطع الليجو التي نحتاجها لملء الفراغ أقل، وفرصة الاختيار في طريقة ملء هذا الفراغ أيضًا أقل، ومساحة الخيال الذي يمكننا استخدامه لاختراع وابتكار أشكال معينة داخل المسافات البينية بالضرورة أيضًا ستكون أقل، وكلما قلت المسافات تقل فرص الاختيار، حتى تصبح المسافة بين قطعتين هي مسافة لقطعة ليجو واحدة، فحتمًا ستكون قطعة بعينها، لا نملك حق اختيار غيرها. هذا ما نعنيه بمتاهة السجل الأحفوري الشبيهة بمكعبات الليجو، حيث نلاحظ أن المساحة الأعظم منه خالية من الأحافير، ما يجعله مجرد سراب آخر، لا يمكننا من الوصول لاستنتاج ذي قيمة من خلاله، حيث لا يوجد أي قيد يمكن اختباره لاختيار مكان تموضع الأحافير بداخله، ولكي يتم تحديد مسار تسكين الأحافير يجب أن يتوافر كم هائل من الأشكال الأحفورية المتدرجة مورفولوجيا، لتقليص هذه المسافات الزمنية، فكلما قلت المسافة الزمنية والشكلية بين أحفورتين كانت فرص التسكين لشكل انتقالي بينهما أكثر صرامة.

نحن نستشعر الآن أننا بالفعل داخل متاهة حقيقية مسماة بالسجل الأحفوري، والسبب هو فداحة الفواصل الزمنية التي تفصل بين الحفريات، حيث لا نستطيع أن نفترض أي علاقة بينها؛ فهناك فروق شاسعة لملايين الأجيال عبر ملايين السنين تفصل بين الحفريات التي يدعي أنها أسلاف للحيتان –كمثال–، لذا فإن الادعاء بأن هذه الحفريات تمثل "الأشكال الانتقالية" هي مجرد فرضية غير مؤسسة في ظل عدم توافر رابط خطي مباشر بينها، متمثل في آلاف الأحافير المتدرجة.
لكن ما يعزز هذا التيه أكثر، هو أن الإشكالية تتعدى فقر السجل الأحفوري في عدد الأحافير، إلى عدم قدرته على تقديم أي بيانات سلوكية أو جزيئية، أو بيانات عن الأنسجة الرخوة التي تساهم في إجراء المقاربات المطلوبة، وهو ما قام O'Leary بدارسته و توثيقه في السجل الأحفوري المعتمد للحيتان، حيث قرب إلينا الفكرة برسم بياني يحدد حجم البيانات التي يمكن أن يدلي بها سجل أحافير الحيتان، ومَثَّلَ البيانات التي لا يمكن جمعها بمنطقة سوداء شاسعة، تمثل الغالبية العظمى من محتوى المساحة البيانية.(38)
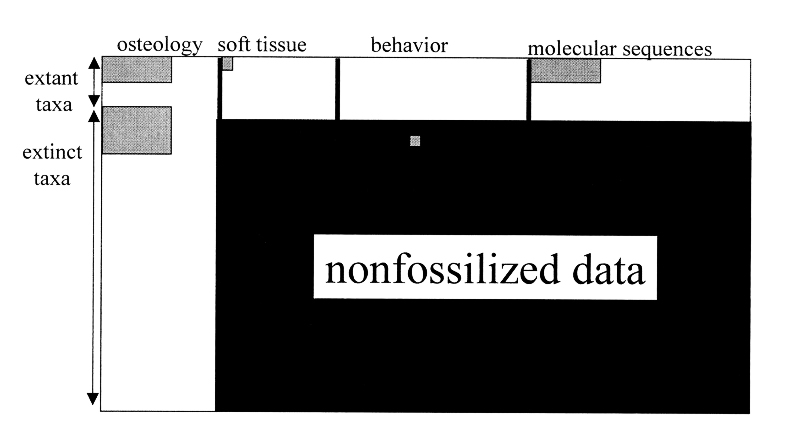
ما سبق يقودنا إلى نتيجة مفادها أنه يمكننا العبث بأريحية تامة داخل هذه الفواصل المورفولوجية والمساحات الزمنية الشاسعة بالسجل الأحفوري، ويمكننا أن نصنع من المعطيات الحقيقية أي شجرة تطورية تحلو لنا وفق عشرات من التوافيق المتاحة بالفعل، ويمكننا أيضًا صنع الأفرع التطورية التي تحلو لنا ، ولا يوجد أي مانع منطقي أو علمي يمنعنا من ذلك مادام أنصار التطور فعلوها بالاعتماد فقط على بعض الخيال، وسأقوم أنا الآن بالبدء بصنع شجرة التطور الخاصة بتصوري لأسلاف الحوت عشوائيًّا، ودون تفكير بمقدمات، لأنني بالتأكيد سأجد دعمًا مماثلًا لما يعتمد عليه أنصار التطور .
يمكنكم تجربة اللعبة فهي سهلة، ويمكنكم اختراع المئات من الأشجار التطورية في المساحة الخالية، ولا يوجد منطق يمكنه إعاقتكم.
فقط يلزمكم بعض الخيال أو الكثير منه، وتبقى الحفريات كقطع لعبة الليجو التي يمكنكم تركيبها وتوظيفها كما تشاؤون لصنع المجسم الذي تريدونه، فبنفس القطع يمكنك أن تصنع سيارة أو بيتا أو سفينة، ويبقى الخيال هو اللاعب الرئيس ما دمنا نواجهه ذلك الفراغ والعوز في الربط بين أي مجموعة بين الحيوانات في هذا السجل بسلف يمكن دعمه بألف سيناريو اعتباطي، لكن عليك أن تتمتع بشيء واحد كما قلنا سابقا؛ الخيال.. والمزيد من الخيال.
هذا الانقطاع والتيه في واقع السجل الأحفوري، وعدم اتساقه مع السيناريو التطوري للحوت، هو ما اعترف به ماكهيدليتسيه G. A. Mchedlidze خبير الحيتان الروسي حين أعرب عن شكوكه الجدية حول ما إذا كانت حفريات مثل باكيسيتوس وأمبيلوسيتوس وغيرهما في سجلات التطور، يمكن اعتبارها أسلاف الحيتان الحديثة، ويرى أنه حتى لو تم قبولها كثدييات مائية، فإنه لا يمكن تمثيلها إلا كمجموعة معزولة تماما عن الحيتان الحديثة.(39)
لكن هذا النموذج الفعلي للسجل الأحفوري –كما تم رصده– الذي نقوم نحن ببناء التخيلات عليه، لو تركناه كما هو بدون أي إضافات أو تخيلات أو عبث، ما الذي يمكننا استقراؤه منه؟
دققوا النظر في الصورة التالية...
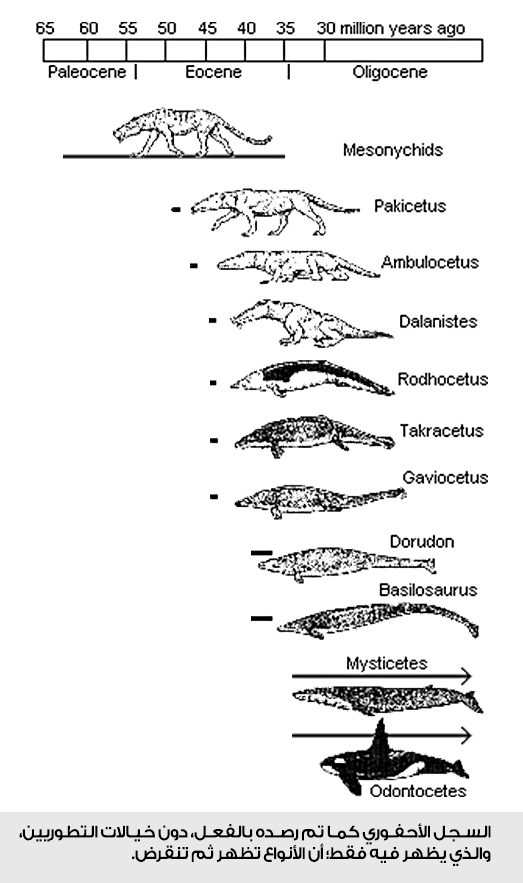
الظهور المفاجئ دليل على الخلق المباشر، وثبات الأنواع دون تغيير ينفي أي تطور حادث. لذلك نصحنا من البداية بعدم الاستدلال بالسجل الأحفوري؛ لأنه دليل مباشر على الخلق.
ملخص ما سبق:
السجل الأحفوري لا يُظهر أي أسلاف مباشرة للحيتان الحالية، لكن أقصى ما يمكنه أن ينبئ به هو وجود أنواع من الحيوانات التي تظهر بعض التشابهات في صفات متفاوتة، هذه الأنواع ظهرت فجأة داخل سجل الأحافير خلال حقب زمنية قديمة، وعاشت لأجيال متعاقبة كما هي دون أن تبدي أي اتجاه نحو التغيير التدريجي والتطور لنوع آخر.
يُظْهِرُ الرصدُ المباشر لسجل الأحافير أن الأنواع المختلفة لمْ تنشأ من خلال التطور من بعضها البعض، بل نشأت بشكل مستقل ومفاجئ، بكل تركيباتها الذاتية. بعبارة أخرى؛ يختلف الخلق من نوع لآخر.
هذه المشاهدة يمكننا رصدها بجلاء على طول السجلات الأحفورية المتاحة لجميع الكائنات الحية دون استثناء، وإن كان يمكننا استقراء أي شيء ذا قيمة من هذه السجلات المتاحة، فإن الاستنتاج المنطقي المباشر والوحيد الذي يمكن أن نستقرئه من تلك البيانات هو الخلق المباشر للأنواع، وعدم قابليتها للتطور.
لنلملم قطع الليجو المتناثرة على الأرض بسرعة قبل أن نتعرض لعقاب الأم الغاضبة، وننتقل إلى لعبة أخرى محيرة لمساعدة الحوت في العثور على أمه، لعبة البحث عن نسب الحوت.
الإشكالية الرابعة: بيانات السجل الأحفوري تتناقض مع البيانات الجزيئية، وإشكالية النسب المجهول

لأكثر من ثلاثة عقود شيد أنصار التطور قلعة من الادعاء لنموذج تطور الحوت من مجموعة الثدييات وسطية الحوافر اللاحمة Mesonychians الشبيهة بالذئاب، بناء على وجود تشابهات في شكل الأسنان وبعض أجزاء الجمجمة، وانصبت كل طاقات البحث في هذا الاتجاه، فأجريت المقاربات ورسمت مخططات عديدة لتأكيد مثل هذا الطريق، وظلت العقيدة التطورية تعتمد بعض هذه المؤشرات المورفولوجية لرسم سجل أحفوري، للإشارة إلى أصل يعود إلى وسطية الحوافر، لكن دائمًا –وكما اعتدنا– في مثل هذه القصص التطورية تأتي الأمواج على قلاع الشاطئ العتيدة، ليتبين أنها لم تكن سوى صروح من الرمال، جرفتها تلك الأمواج في مد المساء حين ارتفع القمر.(40)

في أواخر تسعينيات القرن المنصرم، خرجت بعض القرائن لدراسات استخدمت التقنيات الجزيئية لاستكشاف العلاقات بين مجموعات من الحيوانات، والتي أدلت بنتيجة مغايرة تمامًا للحديث السابق، وادعت أن الحيتان هي الأكثر قرابة لفئة أخرى من الثدييات تضم الماعز والخنازير والأبقار والغزلان والجمال وأفراس النهر، وتدعى مزدوجات الأصابع Artiodactyls.
ذلك من خلال دراسة مقارنات لبعض المناطق غير المكودة من الحمض النووي Noncoding DNA التي أظهرت تشابهًا بين الحوت وأفراس النهرHippopotamusesـ.(41)
البيانات الجزيئية تعارضت بشكل مباشر مع بيانات السجل الأحفوري، حيث إنها جعلت من المقاربات التي تمت بين الحفريات ونسبتها إلى وسطية الحوافرMesonychians –التي يعتقد مسبقا أنها سلف الحيتان بناء على شكل الأسنان وبعض ميزات الجمجمة– لا تؤدي أي دور في التطور، وكتبت العديد من الدوريات العلمية عن مأزق التناقض.(42)
كانت ردة الفعل المتوقعة لعلماء الأحافير المختصين بدراسة تطور الحوت في البداية هي الصدمة، وشككوا في جدية هذه النتائج الجزيئية وقابلوها بالرفض، حتى أن (جينجريتش) أحد أبرز من رسموا مخطط التطور الخاص به اتهم أصحاب هذا الرأي بأنهم مجانين.(43)
في ظل فوضى التناقض بين البيانات، التي لا يمكن دمجها في مخطط تطوري واحد، مما يستلزم رفض أحد طريقي البحث تمامًا، وهذا يمثل إحراجًا لمنهجية التطور؛ لأنه من المفترض –وفقا للداروينية– أن تتسق الدلالات الأحفورية والجزيئية في شجرة فيلوجينية واحدة.
هنا اضطر كثيرون من أنصار التطور المتحمسين لسجل القرابة السابق –لوسطية الحوافر– إلى أكل صنم العجوة الذي ظلوا عليه عاكفين لثلاثة عقود، وقبلوا الأدلة الجزيئية التي تدل على سجل قرابة آخر، لكن ما بال تلك التشابهات في شكل الأسنان والجمجمة الأحفورية التي بنيت عليها قلعة تطور الحوت؟
بالطبع الحل لتلك الإشكالية مُعلب وجاهز للاستهلاك المباشر، وأظن أنكم عرفتموه جيدًا؛ إنه التطور المتقارب convergent evolution، وببساطة تم التراجع عن أوضح سجلات التطور المزعومة، وتم إرجاع التشابه بين تلك الأشكال إلى ما يسمى التطور التقاربي غير المرتبط بسلف، أي أن هذا التشابه في الأسنان وعظام الأذن الذي اعتُمد لرسم القرابة، لم يعد يدل على سلف مشترك كما أدلت الدوريات العلمية.(44)
البيانات الأحفورية غير متناسقة بشكل صارخ مع هذه الفرضية، إذا كانت الدلالة الفيلوجينية للكازين دقيقة فلابد من الاعتراف بفجوات كبيرة في السجل الأحفوري، كذلك انتكاسات مورفولوجية واسعة، وتطور متقارب غير مرتبط بسلف.(45)(46)(47)
من المستغرب هنا، أنه بعد عامين فقط تتغير بوصلة الأدلة الأحفورية، ويبدي كل من (جينجريتش) و(ثويسين) نوعًا من المواربة الذكية، بفتح باب من المواءمة بين مخطط العلاقة الجزيئية واكتشافات حفرية تؤيد الفكرة الجديدة حول قرابة الحيتان من مزدوجات الأصبع artiodactyls .
لكن ما هو الرابط المورفولوجي الذي استخدمه كل من جينجريتش وثويسين لربط مزدوجات الأصبع بالحيتان؟
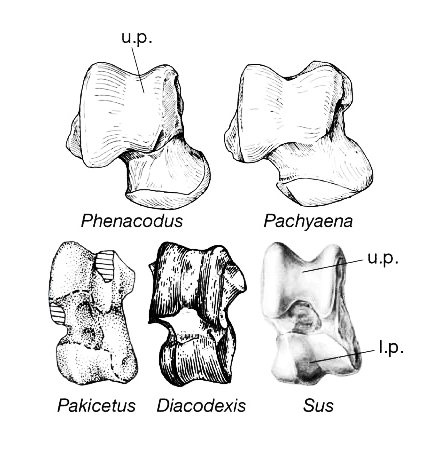
إنها عظمة واحدة فقط!
يمكن أن يكون هذا مثيرا للاستغراب، لكن ماذا لو عرفتم أن هذه العظمة التي تم الاعتماد عليها هي عظمة العقب astragalus إحدى عظام القدم؟
يتساءل أحدكم باستغراب: كيف هذا؟ هل لدى الحوت قدم؟
الإجابة كما اعتمدها علماء الأحافير تكمن في الربط غير المباشر، حيث استخدموا نتيجة البيانات الجزيئية التي اعتبرت الحيتان الأقرب من مزدوجات الأصابع كمقدمة للبحث عن أحافير تمتلك عظامًا مشابهة لتلك التي تميز مزدوجات الأصابع، وكان خيارهم الأمثل هو عظمة العقب التي تشبه البكرة.

بعيدًا عن منطق الاستدلال الدائري هنا، والذي عهدناه على أنصار التطور، يبدو أن جينجرتيش امتلك حظا فائقا أو فانوسًا سحريًّا ليجد للأحفورة رودهوكيتوس التي وجدت بدون أرجل في السابق أرجلًا منفردة، نسبها إلى ذلك النوع، وقال إنها تمتلك عظمة عقب شبيهة بتلك التي في مزدوجات الأصابع.
تزامنا مع ذلك الحظ السحري، يستخرج ثويسين وزملاؤه من مَطْمَرَة عظام في باكستان عظامًا من خلف القحف لحيوان ما ينسبها إلى النوع باكيسيتوس، كما استخرجوا هيكلا عظميًّا لفرد أصغر منه من فصيلة الپاكيسيتيدات، ويسمى Ichthyolestes، وقد امتلك كلاهما عَقِبا يحمل الخصائص المميزة للحافريات الزوجية الأصابع.
لكن الجدل الدائر في أوساط التطور حَوْلَ متاهة النسب، جعلت كلا من جينجريتش وثويسين يتريثان في تأكيد فكرة النسب الجديد، فتركا الباب مفتوحًا باعترافهما عدم دلالة هذه الكشوف على فكرة نسب الحوت الجديد لمزدوجات الأصابع artiodactyls، حيث أظهر جينجريتش أن الأحفورة Rodhocetus تمتلك معالم في أياديها ومعاصمها لا تماثل أي من مزدوجات الأصابع اللاحقة الأخرى. ويُحذر ثويسين من أن البيانات المورفولوجية لا تشير حتى الآن إلى حافريٍّ زوجي الأصابع بعينه –مثل فرس النهر– على أنه أقرب الأقرباء للحيتان، أو أنه يمثل مجموعة شقيقة له. ويقول ثويسين: "لم نتوصل بعد إلى حل لتصنيف الحيتان ضمن الحافريات الزوجية الأصابع، لكنني أظن بأن ذلك سيحدث."(48)(49)
بتأمل بسيط لذلك الحدث المتعلق باكتشاف مفاجئ لأجزاء منفصلة لأحافير (أرجل)، تم نسبتها في السابق لوسطية الحوافر Mesonychians، أو أحافير جديدة بعد تغير الفكرة المسبقة في نسب الحوت، يمكننا أن نخلص إلى فكرة مفادها أن الأحافير يتم توظيفها لدعم تلك الفكرة المسبقة، وليست دلالة حقيقة يتم استقرائها بحيادية، كذلك يمكن التلاعب بها بسهولة لأنها تستند إلى مقارنة بين أجزاء مختارة، وفي كثير من الأحيان يتم تجاهل الاختلافات الأكبر. في أغلب الأحيان تقوم العديد من الأشكال الانتقالية المزعومة على رفات مجزأة لحيوان ما، والتي يمكن توجيهها للعديد من التفسيرات؛ ببساطة يمكننا القول أن سجل الاحافير يتم خلقه ليوافق الدوجما التطورية.
لكن لا يزال السؤال دون إجابة حقيقة؛ من هو السلف القريب من الحيتان، مزدوجات الأصابع Artiodactyls، أم وسطية الحوافر Mesonychians؟
الأدلة المتاحة تضع فرضية تطور الحوت في موقف لا تحسد عليه، خاصة والبيانات لا تظهر أفضلية واضحة نحو أي منهما، فهناك رأي لفريق كبير من أنصار التطور لا زال يدعم فكرة بناء قرابة بين الحيتان ووسطية الحوافر Mesonychians، بناء على شكل الأسنان التي تبرز تشابها. واعتبروا ميزة تشابه عظمة العقب، المميزة لمجموعة مزدوجات الأصابع Artiodactyls، لا تدل على سلف مشترك، إنما نشأت بشكل مستقل convergences homoplasyـ.(50)
هكذا نجد أنفسنا أمام خيارين في الرصد الأحفوري لا يمكن التفضيل بينهما؛ الأول: اعتبار الحيتان أقرب الأقارب لمزدوجات الأصابع Artiodactyls، وتجاهل التشابهات بين الأسنان وشكل الجمجمة، واعتبارها مجرد تقارب convergences.
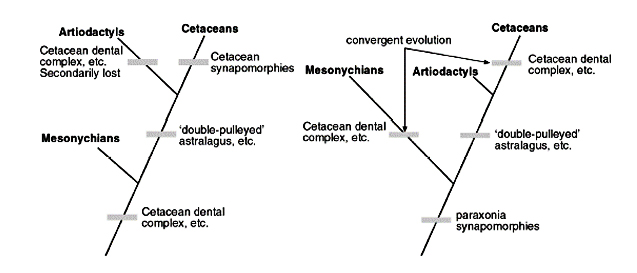
الثاني: اعتبار الحيتان أقرب الأقارب لوسطية الحوافر Mesonychians، وتجاهل التشابهات بين عظمة العقب، واعتبارها مجرد تقارب convergences.
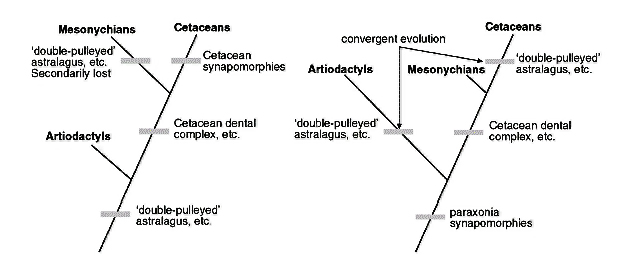
في ظل هذا التضارب والعبث، يمكننا نحن أن نقف باطمئنان موقف المؤيد لكل فريق في نصف موقفه، وهو المتعلق برفض السيناريو الآخر، ونعتبر أن الحوت لا ينتمي لهذا أو ذاك، والقول بعموم أن التقارب convergences أيقونة استدلالية ذكية ضد التطور تهدمه من داخله، فننفي بها دلالة تشابه الأحافير ككل، وننفي بها أيضًا دلالة التشابه الجزيئي الداعم على التطور من أسلاف مشتركة .
حتى البيانات الجزيئية التي من الممكن أن يحتج بها البعض لا يمكن الاعتماد عليها، كما يقر نصير التطور وعالم الأحافير الأشد تعصبًا ضد الخلق (ستيفن جولد Stephen Jay Gould) بتلك الإشكالية وتأثيرها على صحة البيانات، في ظل طمس وانتكاسة شجرة التطور، بسبب التقارب، وأن البيانات المورفولوجية والجزيئية عرضة لمشكلة Homoplasies على حد سواء.(51)
في دراسة أكثر وضوحًا بشأن العلاقة المفترضة بين الثدييات الأرضية والمائية في مجلة Genetics، تم الكشف عن وجود خلاف كبير بين التدابير المورفولوجية والجزيئية، والاعتراف بأن التسلسلات الجزيئية لا تعطينا بالضرورة صورة دقيقة عن النسب.(52)
ملخص ما سبق:
تكشَّف لنا ألعوبة منهجية في طريقة استخدام بعض المعطيات غير الكافية والموهمة، بل والمتضاربة، من قبل أنصار التطور، لصنع قصة رديئة الحبكة، أقل ما يمكن أن توصف به هو العبث والاستهزاء بالعقول.
انتهازية واضحة في الطرح التطوري، هدفها الوصول إلى تحقيق الدوجما المسبقة بأية وسيلة، فحين تتعارض البيانات يتم الاستغناء عن جزء منها بكل بساطة، دون الارتباط بأية منهجية عامة تحكم إطار الاستدلال، ودون مراعاة التناقضات المرصودة في نفس قضية البحث، واللجوء إلى نوع من أسوأ أنواع التدليس العلمي، وهو الانتقاء حسب الحاجة وتجاهل الإشكاليات.
في ظل هذه الفوضى وتوليف الحجج حسب الاستهلاك المحلي دون أي قيد منهجي، فإنه من الواضح جليًّا أننا بالفعل أمام برادايم فكري مسيطر.
الإشكالية الخامسة: معضلة الإطار الزمني لتطور الحوت

يعتقد التطوريون أن الحيتان قد تحولت من السلف الأول الأرضي الكامل باكيسيتوس، الذي عاش قبل 50 مليون سنة، حتى الوصول إلى الحيتان كاملة المعيشة المائية باسيلوصوريات Basilosauridae، التي عاشت قبل 40 مليون سنة، كما تظهر سجلات الأحافير.
بعيدًا عن متاهة النسب المجهول، فإن أي مقترح تطوري لأي حيوان بريّ يمكنه أن يمثل سلفا للحوت، يتطلب اختلافات مورفولوجية هائلة، وتكاليف التحول إلى (الحيتان) باهظة، حيث يجب أن يحدث التطور المزعوم بمعدل عدد مذهل –لا يصدق– من الطفرات (المفيدة) والتكيفات. هذه الفترة الزمنية القصيرة، التي لا تتعدى 10 مليون سنة، والتي هي الإطار الزمني الذي رصد في السجل الأحفوري للتحول من الثدييات الأرضية بالكامل إلى الحيتان المائية بالكامل، شكلت تحديًا لأيّة آلية داروينية يمكن اعتمادها للحصول على هذا الكم الهائل من التحولات الجذرية، فالحيتان لديها العديد من المزايا الفريدة لتمكينها من العيش في الماء.
قطعًا ستحتاج ملامح الهيكل العظمي لتغيير جذري، كذلك الآليات الفسيولوجية –وظائف الأعضاء للكائن الحي–. فعلى سبيل المثال، إنه في وقت مبكر كان من المفترض أن أمبيلوسيتوس يشرب المياه العذبة طوال حياته –قبل 49 مليون سنة–، وكان Protocetid يشرب المياه المالحة –قبل 47 مليون سنة–؛ ما يعني أن تغييرًا متطرفًا في وظائف الأعضاء يجب أن يحدث في فترة لا تزيد عن 3 مليون سنة.
كان سيتعين على Protocetid التحور بطريقة مفيدة لإنتاج التكيفات الفسيولوجية أعلاه خلال هذه الفترة القصيرة جدا، بالإضافة إلى ذلك من المفترض أنها طورت مختلف الآليات الفسيولوجية لتبادل الأكسجين والغطس لمسافات طويلة وتراكم حامض اللبنيك، كذلك تطوير نظام شامل لتخزين الدهون وتنظيم درجة الحرارة في وقت قصير جدًا، وثمة مشكلة أخرى؛ هي اختلاف نوعي تغذية الحيتان والحيوانات البرية وطرق بحثها عن الغذاء، تحتاج الحيتان لتكون جاهزة لهذا أن تكون معدة لممارسة رياضة الغوص العميق، وإمكانية إرضاع صغارها تحت الماء.(53)
واحدة من الإشكاليات العميقة هي أن الثدييات من ذوات الدم الحار، خلقها الله بدرجة حرارة ثابتة للجسم أعلى من الأسماك والزواحف والبرمائيات، والحفاظ على درجة حرارة الجسم الأساسية لكائن ثديي يعيش في محيط من الماء البارد يمثل مشكلة حقيقية. تتغلب الحيتان على تلك الإشكالية بامتلاكها هياكل بيولوجية رائعة ومعقدة تسمى المبادلات الحرارية المعاكسة countercurrent heat exchange للحفاظ على حرارة الجسم الثابتة.(54)
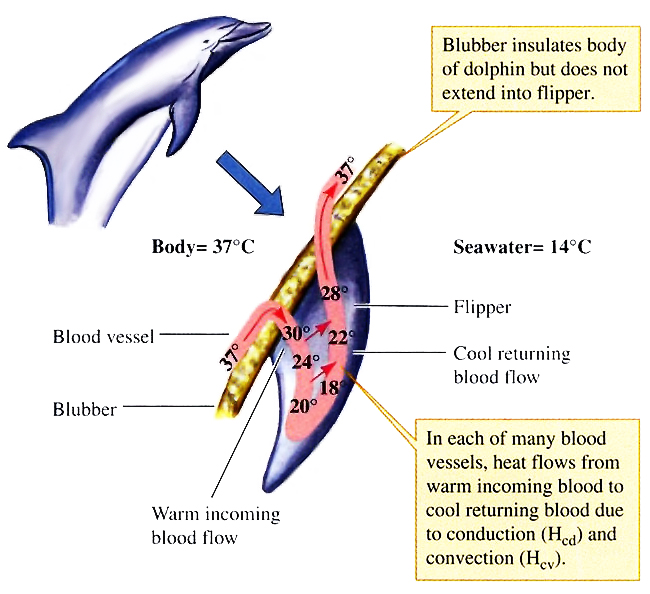
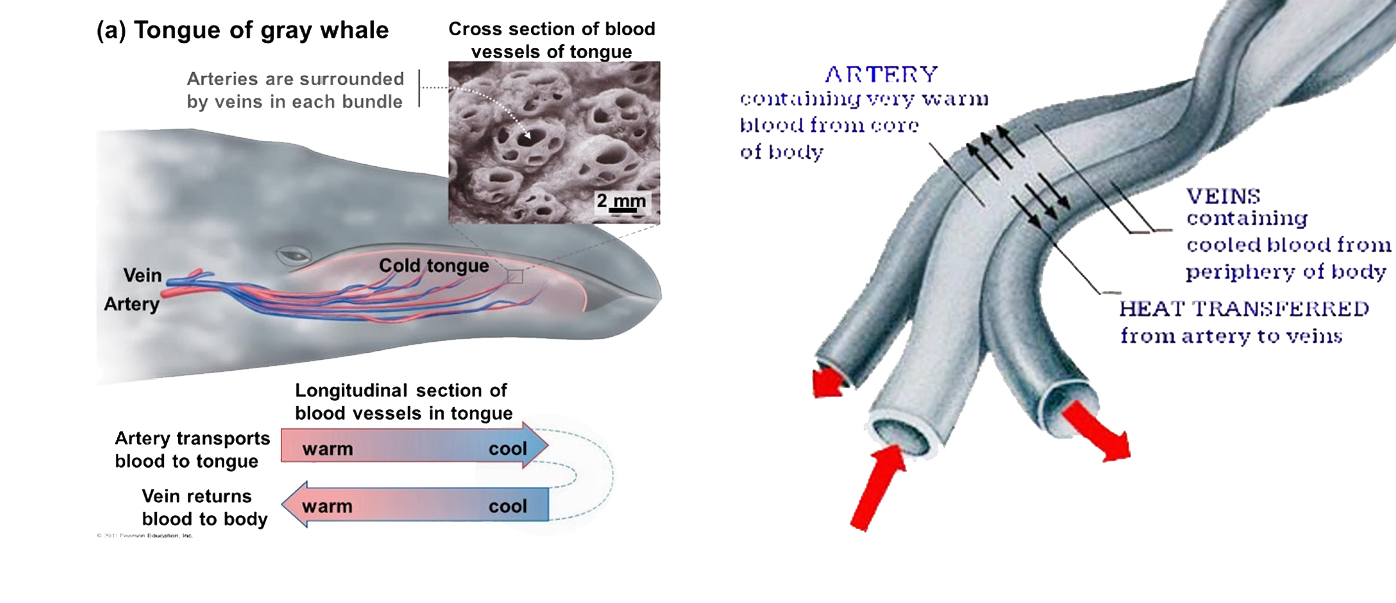
الإشكالية الأكثر تعقيدًا أمام نموذج التطور، هي إمكانية تطوير نظام السونار وتحديد الأماكن بصدى الصوت كوسيلة للاتصال تحت الماء من خلال الموجات الصوتية. والمدهش أن كثيرًا من الحيتانيات لديها هذا النظام الدقيق الذي تحسدها عليه أكثر الغواصات تقدمًا، حيث يمكنها الكشف عن سمكة في حجم كرة الجولف على مسافة سبعين مترًا.
جذبت هذه الآلية العبقرية خبيرًا في نظرية الفوضى يدعى روري هوليت Rory Howlett لدراستها في الدلافين، وتوصل إلى استنتاج مفاده أن تلك الأنماط لابد لها من تصميم رياضي بالغ الدقة لكي تعمل.(55)
هذه الميزة المذهلة لنظام السونار، متمثلة في نتوء دهني على جبين الحيتان والدلافين يسمى البطيخة melon، وهو عبارة عن عدسة متطورة، مصممة لتركيز الموجات الصوتية المنبعثة في شعاع يمكن للحوت أن يوجهه حيث يشاء، هذه العدسة الصوتية هي تجمعات دهنية مختلفة، يجب أن تكون مرتبة في الشكل الصحيح والتسلسل الصحيح، من أجل تركيز أصداء الصوت العائدة، وكل نوع من هذه الدهون فريد ومختلف عن الدهون الطبيعية، فهي تتكون من خلال عملية كيميائية معقدة تتطلب عددًا من الإنزيمات المختلفة.
ولكي يتطور مثل هذا الجهاز يجب على الطفرات العشوائية أن تشكل الإنزيمات الصحيحة لتكوين الدهون الصحيحة، وطفرات أخرى يجب أن تضع هذه الدهون في المكان والترتيب المناسب. إن التطور خطوة بخطوة –تدريجيًّا– لمثل هذا الجهاز ليس ممكنا؛ لأنه إما أن يكون قد تشكل بشكل كامل في المكان والترتيب المناسب، أو أنه سيكون عديم الفائدة، والانتقاء الطبيعي لا يحبذ أشكال وسيطة غير مكتملة، لأنها تمثل عبئا عليه.
لذلك يمكننا أن نعتبر مثل هذه الفسيولوجية من التعقيدات تتحدى آليات التطور؛ لكونها غير قابلة للتدرج أو الاختزال.(56)
وحتى لا نطيل في مئات التفصيلات، فإن ما يُفترض أن يحدث هو تحول شبه كامل لوظائف الأعضاء والملامح التشريحية وتجديد الأسلاك الكهربائية الجينية اللازمة لهذا التحول، وللاختصار نذكر فقط بعض الأمثلة الأخرى التي يجب أن تتطور:
– عيون مصممة لتتمكن من الرؤية بشكل صحيح تحت الماء مع عوامل الانكسار، وتحمل الضغط العالي.
– آذان مصممة بشكل مختلف عن تلك الثدييات البرية، التي تلتقط الموجات الصوتية المحمولة جوًا، مع طبلة أذن محمية من الضغط العالي.
– الجلد يفتقر إلى الشعر والغدد العرقية، وطبقة دهون للعزل الحراري.
– الخياشيم على الجزء العلوي من الرأس (blowholes).
– ذيل الحوت والجهاز العضلي.
– الجنين في موقف المقعدية –للولادة تحت الماء–.
– تعديل الثدي.
– فقدان الحوض والفقرات العجزية.
– إعادة تنظيم الجهاز العضلي.
والآن، بالعودة إلى عنوان الإشكالية، هل الإطار الزمني لتطور الحوت وظهور هذه التحولات المذهلة كافٍ؟
عالم الأحياء التطوري ريتشارد ستيرنبرج Richard Sternberg قام بالاستعانة ببعض آليات التطور المعتمدة لاختبار ذلك الحدث، ووفقا لتلك الحسابات التي أجراها ستيرنبرغ بالاستناد إلى معادلات الوراثة السكانية Population Genetics المطبقة في ورقة للعالمين شميدت Deena Schmidt ودوريت Rick Durrett في مجلة علم الوراثة، فإنه يتوقع حدوث تثبيت لاثنين من الطفرات في إطار زمني يقدر بحوالي ثلاث وأربعين مليون سنة.(57)
بالمقارنة مع حجم هذا التحول المرصود في تطور ثديي بري صغير –شبيه بالغزال أو الذئب أيًا كان التخيل التطوري– إلى حوت ضخم عتيد، فإن تطبيق آليات الوراثة السكانية وتثبيت الطفرات يعتبر هنا أمرًا جنونيّا، ولا يمكن حدوثه في فترة لا تتجاوز عشرة ملايين عام.
لكن ما رأيكم أن نزيد جرعة التعقيد، ونتخطى بها حاجز الجنون إلى الانتحار العقلي التام؛ ففي الآونة الأخيرة تم الإبلاغ عن اكتشاف عظم الفك لأحد الحيتان القديمة في القارة القطبية الجنوبية، بواسطة فريق بحث أرجنتيني، وقالوا إنها لأقدم حوت عاش حياة مائية كاملة قبل تسع وأربعين مليون سنة.(58)
هذا الكشف يدمر تمامًا المخطط الأحفوري السابق، ويُقلص الفترة الزمنية المزعومة لتطور الحوت من عشرة ملايين عام إلى مدة تقل عن ثلاثة ملايين عام أو أقل؛ ليصبح الأمر أقرب للخرافة.
الخاتمة:
لنتكلم بطريقة أكثر حزمًا أمام فاشية الداروينية، تلك التي تدعي زورًا انتهاجها سبيل العلم، ونلخص قصة تطور الحوت في المثال التالي: من المؤكد أن ثمرة الطماطم قد تطورت من عربة المطافئ الحمراء، لكنها فقدت عجلاتها التي كانت تسير عليها يومًا ما، ودليل ذلك أن كليهما أحمر اللون ومملوء بالماء!
لا تسخروا من منطقي وتقولوا وما شأن العجلات باللون، وهل هذا كافٍ لتقرير هذا الاستقراء العجيب؟! فإجابتي ببساطة ستكون إحالتكم لأصدقائنا من أنصار التطور، لأنهم وحدهم من يستطيعون الإجابة عن تلك الإشكالية، وهم وحدهم يستطيعون وضع السيناريو الكامل للتحول الجذري لثمرة الطماطم حتى فقدت عجلاتها من سلفها عربة المطافئ.
وكما قلنا سابقًا؛ الأمر يحتاج فقط للحبكة والمؤثرات، وبعض الخيال... الكثير من الخيال.
المراجع:
(1) Van Valen, “Deltatheridia, A New Order of Mammals,” Bulletin of the American Museum of Natural History 132 (1966): 92.
digitallibrary.amnh.org/dspace/handle/2246/1126
Van Valen, "The Deltatheridia, a new order of mammals." Bull. Am. Mus. Nat. Hist, 1321-126. L. 1966.
icb.oxfordjournals.org/content/41/3/487.full#ref-36
(2) J. G. M. Thewissen and E. M.Williams, "The Early Radiations Of Cetacea (MAMMALIA): Evolutionary Pattern and Developmental Correlations", Annu. Rev. Ecol. Syst. 2002. 33:73–90.
www3.neomed.edu/DEPTS/ANAT/Thewissen/pdf/2002ThewWillAnnRev.pdf
(3) Rebecca Boyle," Jurassic Mammal Fossil Hints At Earlier Split Between Placental Mammals and Marsupials", 08.26.2011 Popsci site.
www.popsci.com/science/article/2011-08/jurassic-mammal-fossil-hints-earlier-mammal-marsupial-split
Zhe-Xi Luo et al, "A Jurassic eutherian mammal and divergence of marsupials and placentals", Nature 476, 442–445 (25 August 2011(.
www.nature.com/nature/journal/v476/n7361/full/nature10291.html
(4) أحمد يحيى، الوحوش الجرابية تهدم أدلة التطور المورفولوجية.
(5) مصادرة على المطلوب، ويكيبيديا العربية.
(6) Ronald R. West , "Paleontology and Uniformitarianism ," in Compass , May 1968, p. 216.
"Contrary to what most scientists write, the fossil record does not support the Darwinian theory of evolution, because it is this theory (there are several) which we use to interpret the fossil record. By doing so, we are guilty of circular reasoning if we then say the fossil record supports this theory".
(7) Gingerich PD, et al. "Origin of Whales in Epicontinental Remnant Seas: New Evidence from the Early Eocene of Pakistan" Science 22 April 1983: Vol. 220.
(8) J. G. M. Thewissen, E. M. Williams, L. J. Roe2 & S. T. Hussain "Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls", Nature 413, 277-281, (20 September 2001).
www.nature.com/nature/journal/v413/n6853/full/413277a0.html
See Also: Christian de Muizon, "Walking with whales", Nature 413, 259-260 (20 September 2001).
www.usca.edu/biogeo/studentinfo/Muizon2001.pdf
"All the postcranial bones indicate that pakicetids were land mammal...
The relatively rigid articulations of the lumbar vertebrae, and the long, slender limb bones— indicate that the animals were runners, moving with only their digits touching the ground".
(9) Douglas H. Chadwick, "Earth’s largest animals are sometimes born with a leg or two, a startling genetic reminder of the time, 50 million years ago, when their ancestors walked on dry land." Nationalgeographic.com.
ngm.nationalgeographic.com/ngm/data/2001/11/01/html/ft_20011101.4.html
(10) G. M. Thewissen, et al, "Fossil evidence for the origin of aquatic locomotion in Archeocete whales," Science, 1994, Vol 263, p. 210–212.
www.sciencemag.org/content/263/5144/210
(11) Steering Committee on Science and Creationism, "Science and Creationism: A View from the National Academy of Sciences", National Academy of Sciences 1999. p.20.
www.nap.edu/openbook.php?record_id=6024&page=20
(12) op. cit (9).
(13) Carroll, "Patterns and Processes of Vertebrate Evolution", Cambridge University Press, p. 335.
(14) Working Group on Teaching Evolution, "Teaching About Evolution and the Nature of Science", National Academy of Sciences 1998, p.18.
www.nap.edu/openbook.php?record_id=5787&page=18
“"I speculated that it might have had a fluke; I now doubt that Rodhocetus would have had a fluked tail.”
(15) Carl Werner, "Evolution: the Grand Experiment" Vol. 1, p.143.
www.thegrandexperiment.com
“Since then we have found the forelimbs, the hands, and the front arms of Rodhocetus, and we understand that it doesn’t have the kind of arms that can spread out like flippers on a whale.”
(16) Whale evolution - why the deception? – Youtube.
www.youtube.com/watch?v=5G5vAc5_VJo
(17) Gingrich and others (1994) published this reconstruction of the skeleton of Rodhocetus kasrani.
taxonomy.zoology.gla.ac.uk/~rdmp1c/teaching/l1/evolution/l1/geology.html
(18) op. cit (14).
(19) Whale Evolution? - Exposing The Deception In The Fossil Record - Dr. Terry Mortenson – Metacafe.
www.metacafe.com/watch/4032568/whale_evolution_exposing_the_deception_in_the_fossil_record_dr_terry_mortenson/
(20) Darwin "The Origin of Species", Crown Publishers, New York, 1979, p. 292
"The number of intermediate varieties, which have formerly existed on the earth, (must) be truly enormous. Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely graduated organic chain; and this, perhaps, is the most obvious and gravest objection which can be urged against my theory".
(21) David Raup, "Conflicts between Darwin and Paleontology , Field Museum of Natural History Bulletin" , Vol. 50, No. 1, 1979 , p. 22
"We are now about 120 years after Darwin and the knowledge of the fossil record has been greatly expanded. We now have a quarter of a million fossil species, but the situation hasn't changed much. The record of evolution is still surprisingly jerky and, ironically, we have even fewer examples of evolutionary transition than we had in Darwin's time. By this I mean that some of the classic cases of Darwinian change in the fossil record, such as the evolution of the horse in North America, have had to be discarded or modified as a result of more detailed information".
(22) Duane T. Gish, "The Origin of Mammals in Creation: The Cutting Edge" (1982) , p. 76
"If this view of evolution is true, the fossil record should produce an enormous number of transitional forms. Natural history museums should be overflowing with undoubted intermediate forms. About 250,000 fossil species have been collected and classified. These fossil species have been collected at random from rocks that are supposed to represent all of the geological periods of earth's history. Applying evolution theory and the laws of probability, most of these 250,000 species should represent transitional forms. Thus, if evolution is true, there should be no doubt, question, or debate as to the fact of evolution".
(23) Phyletic gradualism, The Veritas Forum.
www.veritasucsb.org/library/origins/CATALOG/FIGE.html
(24) Eldredge, N., "Macro-Evolutionary Dynamics : Species , Niches, and Adaptive Peaks", McGraw-Hill Publishing Company 1989 , New York , p. 22
"Most families, orders, classes, and phyla appear rather suddenly in the fossil record, often without anatomically intermediate forms smoothly interlinking evolutionarily derived descendant taxa with their presumed ancestors".
(25) G.G. Simpson, "in The Evolution of Life", p. 149.
"It is a feature of the known fossil record that most taxa appear abruptly. They are not, as a rule, led up to by a sequence of almost imperceptible changing forerunners such as Darwin believed should be usual in evolution".
(26) A.H. Clark, "The New Evolution: Zoogenesis" , p. 114.
"All the major groups of animals have maintained the same relationship to each other from the very first [from the very lowest level of the geologic column]. Crustaceans have always been crustaceans, echinoderms have always been echinoderms, and mollusks have always been mollusks. There is not the slightest evidence which supports any other viewpoint".
(27) James W. Valentine and Cathryn A. Campbell, "Genetic Regulation and the Fossil Record , " American Scientist , Vol. 63 , November , 1975, p. 673
"The abrupt appearance of higher taxa in the fossil record has been a perennial puzzle. Not only do characteristic and distinctive remains of phyla appear suddenly, without known ancestors, but several classes of phylum, orders of a class, and so on, commonly appear at approximately the same time, without known intermediates".
(28) Kemp, Tom "A Fresh Look at the Fossil Record" , New Scientist, Vol. 108, No. 1485, December 5, 1985) , p. 66
"As is now well known, most fossil species appear instantaneously in the fossil record".
(29) Eldredge, N. et al "The Myths of Human Evolution" Columbia University Press; 1982, p. 59.
"The record jumps, and all the evidence shows that the record is real: the gaps we see reflect real events in life's history – not the artifact of a poor fossil record".
(30) Stanley, S. M., 1981 "The New Evolutionary Timetable: Fossils, Genes, and the Origin of Species" Basic Books Inc. Publishers N.Y., p.3.
"For more than a century biologists have portrayed the evolution of life as a gradual unfolding ... Today the fossil record ... is forcing us to revise this conventional view".
(31) opt cit (29) p. 163.
"The fossil record flatly fails to substantiate this expectation of finely graded change".
(32) Gould , S.J. "Evolution's Erratic Pace", Natural History, vol. 86, May 1977.
"The history of most fossil species include two features particularly inconsistent with gradualism:
1) Stasis – most species exhibit no directional change during their tenure on earth. They appear in the fossil record looking much the same as when they disappear; morphological change is usually limited and directionless;
2) Sudden appearance – in any local area, a species does not arise gradually by the steady transformation of its ancestors; it appears all at once and 'fully formed'".
(33) opt cit (23).
(34) Ibid.
(35) Ricklefs, Robert E., “Paleontologists Confronting Macroevolution,” Science, vol. 199, 1978, p.59.
"The Eldredge–Gould concept of punctuated equilibria has gained wide acceptance among paleontologists. It attempts to account for the following paradox: Within continuously sampled lineages, one rarely finds the gradual morphological trends predicted by Darwinian evolution; rather, change occurs with the sudden appearance of new, well–differentiated species. Eldredge and Gould equate such appearances with speciation, although the details of these events are not preserved. …The punctuated equilibrium model has been widely accepted, not because it has a compelling theoretical basis but because it appears to resolve a dilemma. Apart from the obvious sampling problems inherent to the observations that stimulated the model, and apart from its intrinsic circularity (one could argue that speciation can occur only when phyletic change is rapid, not vice versa), the model is more ad hoc explanation than theory, and it rests on shaky ground".
(36) Ronald R. West , "Paleontology and Uniformitarianism" Compass , May 1968, p. 216.
"Contrary to what most scientists write, the fossil record does not support the Darwinian theory of evolution, because it is this theory (there are several) which we use to interpret the fossil record. By doing so, we are guilty of circular reasoning if we then say the fossil record supports this theory".
(37) Mark Ridley , "Who doubts evolution?", New Scientist, vol. 90, 25 June 1981 , p. 831
"In any case, no real evolutionist, whether gradualist or punctuationist, uses the fossil record as evidence in favour of the theory of evolution as opposed to special creation".
(38) The Phylogenetic Position of Cetaceans: Further Combined Data Analyses, Comparisons with the Stratigraphic Record and a Discussion of Character Optimization - Oxford Journals.
icb.oxfordjournals.org/content/41/3/487/F9.expansion.html
(39) G. A. Mchedlidze, "General Features of the Paleobiological Evolution of Cetacea" A. A. Balkema 1984.
onlinelibrary.wiley.com/doi/10.1002/iroh.19860710425/abstract
(40) Fig: Ectotympanic bones of Pakicetus and the modern dolphin Lagenorhynchus. This bone surrounds the middle ear cavity like a bowl. In all cetaceans, the medial wall of the ectotympanic is very thick, as indicated by the white line, and is called the involucrum.
www.springerimages.com/Images/LifeSciences/1-10.1007_s12052-009-0135-2-11
(41) Mitsuru Shimamura. et al, " Molecular evidence from retroposons that whales form a clade within even-toed ungulates", Nature 388, 666-670 (14 August 1997).
www.nature.com/nature/journal/v388/n6643/full/388666a0.html
(42) Fig: Cladograms depicting competing phylogenetic hypotheses for the position of cetaceans among ungulates. (A) traditional hypothesis of relationships with a monophyletic Artiodactyla containing two major clades and the extinct (†) clade Mesonychia is the sister taxon of cetaceans; also the hypothesis supported by data that fossilize (i.e., osteological and dental data, see O'Leary, 1999); and (B) the cladogram supported by molecular data (e.g., Gatesy et al., 1999a), “Artiodactyla” is paraphyletic. Molecular sequence data alone cannot inform on the position of the wholly extinct clade Mesonychia.
icb.oxfordjournals.org/content/41/3/487/F1.expansion.html
Kate Wong, "The Mammals That Conquered the Seas: New fossils and DNA analyses elucidate the remarkable evolutionary history of whales", SCIENTIFIC AMERICAN MAY 2002.
www.miracosta.edu/home/kmeldahl/articles/whaleevol.pdf
Maureen A. O'Leary, "The Phylogenetic Position of Cetaceans: Further Combined Data Analyses, Comparisons with the Stratigraphic Record and a Discussion of Character Optimization", Amer. Zool. (2001) 41 (3): 487-506.
icb.oxfordjournals.org/content/41/3/487.full
Gavin J. P. Naylor, "Are The Fossil Data Really At Odds With The Molecular Data? Morphological Evidence For Cetartiodactyla Phylogeny Reexamined", Syst. Biol. 50(3):444 – 453, 2001.
Abstract: www.ncbi.nlm.nih.gov/pubmed/14668123
Full Article: www.naylorlab.scs.fsu.edu/Publications/Cetartiodactyla.pdf
Full Article(alt): sysbio.oxfordjournals.org/content/50/3/444.full.pdf
University Of Michigan. "New Fossils Suggest Whales And Hippos Are Close Kin". ScienceDaily, 20 September 2001.
www.sciencedaily.com/releases/2001/09/010920072245.htm
Jean-Renaud Boisseri et al, "The position of Hippopotamidae within Cetartiodactyla", PNAS vol. 102 no. 5
www.pnas.org/content/102/5/1537
(43) Ibid p.78.
"The whale-hippo connection did not sit well with paleontologists. “I thought they were nuts,” Gingerich recollects. “Everything we’d found was consistent with a mesonychid origin. I was happy with that and happy with a connection through mesonychids to artiodactyls.” Whereas mesonychids appeared at the right time, in the right place and in the right form to be considered whale progenitors, the fossil record did not seem to contain a temporally, geographically and morphologically plausible artiodactyl ancestor for whales, never mind one linking whales and hippos specifically.."
(44) J Gatesy. et al, "Evidence from milk casein genes that cetaceans are close relatives of hippopotamid artiodactyls." Mol Biol Evol (1996) 13 (7): 954-963.
mbe.oxfordjournals.org/content/13/7/954
(45) Ibid.
"Paleontological information is grossly inconsistent with this hypothesis. If the casein phylogeny is accurate, large gaps in the fossil record as well as extensive morphological reversals and convergences must be acknowledged".
(46) Gavin J. P. Naylor, "Are The Fossil Data Really At Odds With The Molecular Data? Morphological Evidence For Cetartiodactyla Phylogeny Reexamined", Syst. Biol. 50(3):444 – 453, 2001.
"The second more speculative hypothesis is that mammalian teeth are more evolutionarily plastic than was originally believed, an that any phylogenetic signal initially present in the dental data has been eroded because of convergent evolution".
(47) obt cit (8).
"Cetaceans and some Mesonychians have dental similarities and an elongated skull, but these features are probably the result of convergent evolution".
(48) Kate Wong, "The Mammals That Conquered The Seas: New Fossils And Dna Analyses Elucidate The Remarkable Evolutionary History Of Whales", Scientific American May 2002.
"Gingerich notes that Rodhocetus and anthracotheres share features in their hands and wrists not seen in any other later artiodactyls. Thewissen agrees that the hippo hypothesis holds much more appeal than it once did. But he cautions that the morphological data do not yet point to a particular artiodactyl, such as the hippo, being the whale’s closest relative, or sister group. “We don’t have the resolution yet to get them there,” he remarks, “but I think that will come".
(49) Milankovitch , et al, "Even-toed fingerprints on whale ancestry", Nature 388:623, 1997.
faculty.virginia.edu/bio202/202-2002/Lectures%2020202/thewissen%20et%20al%201997.pdf
"then either Mesonychians are not closely related to cetaceans (and many dental characters are convergent), or the specialized heel morphology is not the exclusive character that many morphologists take it to be. It may have evolved several times independently in artiodactyls, or have been lost in the Mesonychians/Cetacean clade. The complete astragalus of an early cetacean would probably shed light on this issue".
(50) Naylor, G.J.P. and Adams, D.C. "Are the fossil data really at odds with the molecular data? Morphological evidence for cetartiodactyla phylogeny reexamined", Systematic Biology 50(3):444–453, 2001
sysbio.oxfordjournals.org/content/50/3/367.full.pdf
Matthee et al., "Mining the mammalian genome for artiodactyl systematics", Systematic Biology 50 (3 ): 388, 2001.
Milankovitch et al. , "Cetaceans are highly derived artiodactyls", Thewissen, Ref. 35, p. 127
(51) Zhexi Luo, "Evolution: In search of the whales' sisters", Nature, Vol. 404, No. 6775, p. 235-239 (1998).
www.stephenjaygould.org/ctrl/news/file017.html
"Both morphological and molecular data are vulnerable to the problem of homoplasies — reversals to ancestral conditions or parallel changes in different lineages that can camouflage the true phylogeny. In this sense, neither approach is better than the other. For instance, the ear region of the skull, traditionally considered to be a good source of highly stable characters, shows some glaring homoplasies among the ungulates and cetaceans 4, 5. Moreover, the fossil record of many early divergent fossil taxa is incomplete, resulting in ambiguities in morphological estimates".
(52) Janke A, et al, "The marsupial mitochondrial genome and the evolution of placental mammals." Genetics 137:243-256.
Abstract : www.ncbi.nlm.nih.gov/pubmed/8056314
Full Pdf : www.genetics.org/content/137/1/243.full.pdf
"The entire mitochondrial genome of the American opossum has been sequenced. Two major differences with placental genomes are noted. First, the sequence of five tRNA genes is different. Second, the aspartic acid tRNA has an anticodon not normally found in the mitochondrion. Eight of thirteen mitochondrial genes are said to exhibit clocklike divergence rates. Lineage divergences based on these genes and calibrated against the geologic time–scale indicate a date of 35 Ma for the divergence of the closely related rat and mouse, compared with 41 Ma for divergence of cow and whale.
These results reveal a large discordance between morphological and molecular measures of similarity. Rats and mice are classified in the same Family, while cows and whales are classified in different Orders. Perhaps molecular sequences are not necessarily giving us an accurate picture of ancestry".
(53) J. G. M. Thewissen, "Evolution of cetacean osmoregulation", Nature 381, 379 - 380 (30 May 1996).
www.nature.com/nature/journal/v381/n6581/abs/381379b0.html
(54) P. F. Scholander et al, "Countercurrent Heat Exchange and Vascular Bundles in Sloths", Journal of Applied Physiology May 1, 1957 vol. 10 no. 3 405 - 411.
jap.physiology.org/content/10/3/405.short
Fig: Aquatic birds and mammals, such as penguins, seals, and whales, can be endothermic in an aquatic environment for two major reasons: First, they are all air breathers and do not expose a large respiratory surface to the surrounding water. Second, many endothermic aquatic animals, including penguins, seals, and whales, are well insulated from the heat-sapping external environment by a thick layer of fat, while others, such as the sea otter, are insulated by a layer of fur that traps air. The parts of these animals that are not well insulated, principally appendages, are outfitted with countercurrent heat exchangers, vascular structures that reduce the rate of heat loss to the surrounding aquatic environment.
biology-forums.com/index.php?action=gallery;sa=view;id=1642
Heyning JE, Mead JG., "Thermoregulation in the mouths of feeding gray whales.", Science. 1997 Nov 7;278(5340):1138-9.
www.ncbi.nlm.nih.gov/pubmed/9353198
(55) Rory Howlett, "Flipper's secret.", NewScientist (28 June 1997).
www.newscientist.com/article/mg15420884.700-flippers-secret.html
(56) Usha Varanasi. et al, "Molecular basis for formation of lipid sound lens in echolocating cetaceans", Nature 255, 340 - 343 (22 May 1975).
www.nature.com/nature/journal/v255/n5506/abs/255340a0.html
Capt. David Williams, "Loss Of Navigation In Beached Whales And Dolphins", Deafwhale site.
deafwhale.com/why_whales_beach/navigation_failure.htm
(57) Durrett R, Schmidt D."Waiting for two mutations: with applications to regulatory sequence evolution and the limits of Darwinian evolution", Genetics, 2008 Nov;180(3):1501-9.
www.ncbi.nlm.nih.gov/pubmed/18791261
Whale Evolution Vs. Population Genetics - Richard Sternberg PhD. in Evolutionary Biology – Metacafe.
www.metacafe.com/watch/4165203/whale_evolution_vs_population_genetics_richard_sternberg_phd_in_evolutionary_biology/
(58) Michael Warren, "Ancient whale jawbone found in Antarctica", NBC News 10/11/2011.
www.nbcnews.com/id/44867222/ns/technology_and_science-science/
-
الثلاثاء PM 02:01
2021-11-16 - 1590